آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Counting Residues |


|
|
Read More
Date: 6-11-2020
Date: 16-6-2021
Date: 7-4-2021
|
Counting Residues
Natural proteins have integral numbers of each of the twenty amino acids in a polypeptide chain, but most currently accepted methods of determining this number by amino acid analysis yield only a nonintegral ratio of moles of amino acid per mole of protein. This value is rarely close to an integer because of experimental error and uncertainty about the molecular weight of the protein. The method described here determines the integral number of residues of certain amino acids, independently of any other property of the polypeptide chain, including its molecular weight. Then this information is combined with amino acid analysis to determine a more accurate value for the molecular weight and the number of other amino acids. The method also provides information about the net charge on a protein.
The general procedure is to modify the N residues of a given amino acid gradually and specifically, so as to generate a complete spectrum of molecules with 0, 1, 2, 3, ¼, N of the groups modified. A single modification reaction can be used, varying the average extent of the reaction. Alternatively, one uses the competition with a related, but distinguishable reagent and carry out the modification to completion. Then the modified species generated are counted after separating them by a technique sensitive only to the number of groups modified by the particular reagent. The most useful modifications are those that alter the electric charge of a residue. Then the number of modifications introduced is determined by separating the various species by electrophoresis, isoelectric focusing, or ion exchange chromatography. The separation must be sensitive only to the number of groups modified in charge, not which particular ones in the polypeptide chain. This is usually accomplished by carrying out the modification and the separation on unfolded molecules, where differences between the N residues are minimized. There should be (N + 1) species present.
The thiol groups of cysteine residues are reacted with a reagent like iodoacetic acid, which introduces an acidic group. This reaction competes with that with the closely related iodoacetamide, which is neutral and does not introduce a charged group. The reaction of the thiol groups is taken to completion, and the number of acidic groups introduced is varied by varying the ratio of iodoacetic acid to iodoacetamide (Fig. 1). The number of acidic groups introduced is determined by electrophoresis under denaturing conditions. In addition to the number of total cysteine residues or thiol groups, the number of disulfide bonds can be determined by counting the thiol groups present before and after reduction of the disulfide bonds.

Figure 1. Counting the six cysteine residues of reduced BPTI. The reduced protein was reacted with the neutral iodoacetamide (1st lane), acidic iodoacetic acid (5th lane), and mixtures of the two in the ratios 1:1, 1:3, and 1:9 in lanes 2, 3, and 4, respectively. Lane 6 contains a mixture of equal portions of the samples applied to lanes 1 to 5. Electrophoresis of the basic protein was in 8 M urea from top to bottom. The electrophoretic mobility is decreased in proportion to the number of acidic carboxymethyl groups on each molecule; the number for each of the electrophoretic bands is indicated at the right. The competition between the two reagents indicates that iodoacetamide reacts three times more rapidly than iodoacetic acid under the conditions used here.
The amino groups of lysine residues and at the N-terminus can be modified specifically by reacting them with a reagent like succinic anhydride, which replaces a basic amino group with an acidic group. Consequently, the net charge of the protein can change by up two units for each amino group reacted. The reaction is varied in its extent by varying the amount of reagent added to the unfolded protein (Fig. 2).
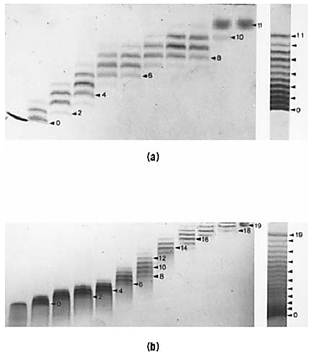
Figure 2. Counting amino groups of bovine ribonuclease A (a) and horse ferricytochrome c (b) by electrophoretically separating the mixtures produced by progressive succinylation. The original unmodified protein is on the left, and the degree of succinylation increases to the right. Electrophoresis was in 8 M urea at pH 3.6 from top to bottom. The separate lane on the far right is of a mixture obtained by combining all of the individual samples. In this case, electrophoresis was at the slightly lower pH of 3.45. Alternating bands are marked by arrows, and the number of succinyl groups is indicated for a few of the bands. The results confirm the presence of 11 and 19 amino groups in ribonuclease A and cytochrome c, respectively.
Other amino acids for which specific modifications are possible may be counted in the same way, in theory, but the specific techniques have not yet been developed.
In each of the previous procedures, the number of groups modified is counted by the number of new bands apparent by electrophoresis. Electrophoretic mobility is proportional to the net charge of the protein molecule, which is changed gradually by modifiying specific residues. Therefore, the net charge on the original protein molecule can be determined by measuring the degree of modification necessary to give the protein zero electrophoretic mobility or by using stepwise modification to
establish a scale for electrophoretic mobility as a function of changes in net charge.
References
1. T. E. Creighton (1980) Nature 284, 487–489.
2. M. Hollecker and T. E. Creighton (1980) FEBS Lett. 119, 187–189.



|
|
|
|
اكتشاف تأثير صحي مزدوج لتلوث الهواء على البالغين في منتصف العمر
|
|
|
|
|
|
|
زهور برية شائعة لتر ميم الأعصاب التالفة
|
|
|
|
|
|
جمعيّة العميد وقسم الشؤون الفكريّة تدعوان الباحثين للمشاركة في الملتقى العلمي الوطني الأوّل
|
|
|
|
الأمين العام المساعد لجامعة الدول العربية السابق: جناح جمعية العميد في معرض تونس ثمين بإصداراته
|
|
|
|
المجمع العلمي يستأنف فعاليات محفل منابر النور في واسط
|
|
|
|
برعاية العتبة العباسيّة المقدّسة فرقة العبّاس (عليه السلام) تُقيم معرضًا يوثّق انتصاراتها في قرية البشير بمحافظة كركوك
|