























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Telomeres Are Synthesized by a Ribonucleoprotein Enzyme
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 24-3-2021
24-3-2021
 2675
2675




+
-
20
Telomeres Are Synthesized by a Ribonucleoprotein Enzyme
KEY CONCEPTS
-Telomerase uses the 3′–OH of the G+T telomeric strand and its own RNA template to iteratively add tandem repeats (5′-TTAGGG-3′ in humans) to the 3′ end at each chromosomal terminus.
- Telomerase uses a reverse transcriptase to extend the very ends of the chromosomes and solve the so-called end replication problem.
The telomere has three widely conserved functions:
- The first is to protect the chromosome end. Any other DNA end —for example, the end generated by a double-strand break—becomes a target for repair systems. The cell must be able to distinguish the telomere.
- The second is to allow the telomere to be extended. If it is not extended, it becomes shorter with each replication cycle (because replication cannot initiate at the very end).
- The third is to facilitate meiotic chromosome reorganization for efficient pairing and recombination of homologous chromosomes.
Proteins that bind to the telomeres contribute to the solution of all of these. In yeast, different sets of proteins solve the first two problems, but both are bound to the telomere via the same protein, Cdc13:
- The Stn1 protein protects against degradation (specifically, against any extension of the degradation of the C-A strand that generates the G-tail).
- A telomerase enzyme extends the C-A–rich strand. Its activity is influenced by two proteins that have ancillary roles such as controlling the length of the extension.
The telomerase uses the 3′–OH of the G+T telomeric strand as a primer for synthesis of tandem TTGGGG repeats. Only dGTP and dTTP are needed for the activity. The telomerase is a large ribonucleoprotein that consists of a templating RNA (encoded by TLC1 in yeast, hTERC in humans) and a protein with catalytic activity (encoded by EST2 in yeast, hTERT in humans). The RNA component is typically short (159 bases long in Tetrahymena, and 451 bases long in humans, though 1.3 kb in yeast) and includes a sequence of 15 to 22 bases that is identical to two repeats of the C-rich repeating sequence. This RNA provides the template for synthesizing the G-rich repeating sequence. The protein component of the telomerase is a catalytic subunit that can act only upon the RNA template provided by the nucleic acid component.
FIGURE 1 shows the action of telomerase. The enzyme progresses discontinuously: The template RNA is positioned on the DNA primer, several nucleotides are added to the primer, and then the enzyme translocates to begin again. The telomerase is a specialized example of a reverse transcriptase, an enzyme that synthesizes a DNA sequence using an RNA template . We do not know how the complementary (C-A–rich) strand of the telomere is assembled, but we can speculate that it could be synthesized by using the 3′–OH of a terminal G-T hairpin as a primer for DNA synthesis.
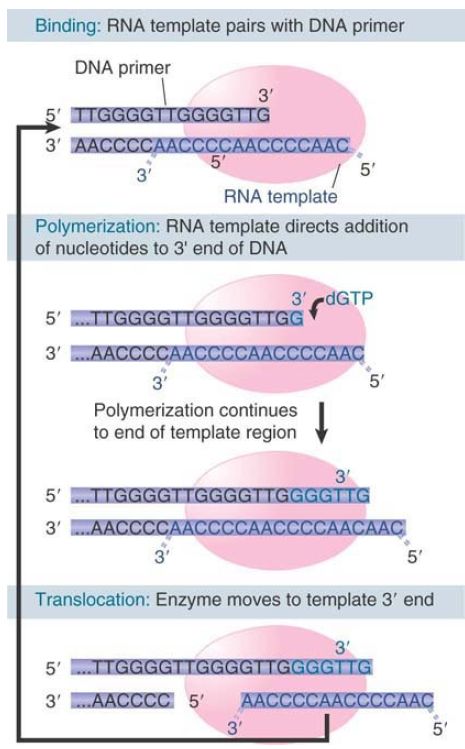
FIGURE 1. Telomerase positions itself by base pairing between the RNA template and the protruding single-stranded DNA primer. It adds G and T bases, one at a time to the primer, as directed by the template. The cycle starts again when one repeating unit has been added.
Telomerase synthesizes the individual repeats that are added to the chromosome ends, but does not itself control the number of repeats. Other proteins are involved in determining the length of the telomere. Some have been identified by the est1 and est3 mutants in yeast, which have altered telomere lengths. These proteins bind telomerase and can influence the length of the telomere by controlling the access of telomerase to its substrate. Researchers have identified proteins that bind telomeres in mammalian cells, including homologs of EST1, but less is known about their functions.
Each organism has a characteristic range of telomere lengths. They are long in mammals (typically 5 to 15 kb in humans) and short in yeast (typically around 300 bp in S. cerevisiae). The basic control mechanism is that the probability that a telomere will be a substrate for telomerase increases as the length of the telomere shortens; we do not know if this is a continuous effect or if it depends on the length falling below some critical value. When telomerase acts on a telomere, it can add several repeating units.
The enzyme’s intrinsic mode of action is to dissociate after adding one repeat; addition of several repeating units depends on other proteins that cause telomerase to undertake more than one round of extension. The number of repeats that is added is not influenced by the length of the telomere itself, but instead is controlled by ancillary proteins that associate with telomerase.
The minimum features required for existence as a chromosome are as follows:
-Telomeres to ensure survival
-A centromere to support segregation
-An origin to initiate replication
All of these elements have been put together to construct a yeast artificial chromosome (YAC; see the chapter titled Methods in Molecular Biology and Genetic Engineering). This is a useful
method for perpetuating large sequences. It turns out that the synthetic chromosome is stable only if it is longer than 20 to 50 kb.
We do not know the basis for this effect, but the ability to construct a synthetic chromosome allows us to investigate the nature of the segregation device in a controlled environment.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















