آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Asexual Reproduction of Flowes |


|
|
Read More
Date: 29-10-2015
Date: 23-10-2016
Date: 19-10-2016
|
Asexual Reproduction of Flowes
Sexual reproduction in flowering plants involves the flower itself, which produces the necessary cells and structures. To understand flower structure, one must first understand:
THE PLANT LIFE CYCLE
The life cycle of mammals such as humans is simple: Diploid adults have sex organs that produce haploid sex cells called gametes, either sperms or eggs, by meiosis. Individuals that produce sperms are called males, of course, and individuals that produce eggs are females. One sperm and one egg are brought together forming a new single diploid cell, the fertilized egg or zygote, which then grows to become a new individual that is diploid and resembles its parents.
In plants, the life cycle is more complex. The plants you are familiar with—trees, shrubs, and herbs—are all just one phase of the plant life cycle, called the sporophyte phase or sporophyte generation. A critical factor is that sporophytes are always diploid, like most adult animals, and they have sex organs (located in the flowers in angiosperms) with cells that are capable of undergoing meiosis. In animals, meiosis results in haploid gametes, but in plants it results in haploid spores (Fig. 1). The difference between gametes and spores is great: Gametes can fuse with other gametes in a process called syngamy or fertilization, thereby producing the diploid zygote. A gamete that does not undergo syngamy dies because it cannot live by itself and usually cannot grow into a new, haploid individual.
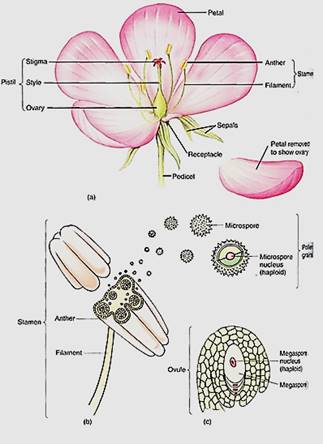
FIGURE 1: (a) Most flowers have four types of structures: (1) sepals that typically protect the young flower, (2) petals that usually attract pollinators, (3) stamens that produce pollen, which produce sperm cells, and (4) a pistil whose ovary is involved in producing egg cells (the pistil shown here is actually composed of five parts called carpels which have merged into one structure). Whereas no one mammal ever has both sperm- producing organs and egg-producing organs, almost all plants do have two types of spore- producing organs, the stamens and the carpels. (b) In the anthers of stamens, the central cells undergo meiosis and each produces four daughter cells called microspores or pollen daughter cells called microspores or pollen grains. (c) In the flower's central organs, the carpels, are ovules, each containing only one cell that undergoes meiosis; often three of the daughter cells die and the one survivor becomes the megaspore.
Plant spores are just the opposite: They cannot undergo syngamy but they do undergo mitosis and grow into an entire new, haploid plant called a gametophyte. It is called a gametophyte because it is the plant (-phyte) that produces the gametes (gameto-). During sexual reproduction, when a sporophyte reproduces, it does not produce a new diploid plant like itself but rather a haploid plant. Furthermore, in all vascular plants, the haploid gametophyte does not even remotely resemble the diploid sporophyte. It is a tiny mass of cells with no roots, stems, leaves, or vascular tissues, but it is an entire plant (Fig. 2). Gametes are formed by the haploid plants by mitosis, not meiosis. The gametes then undergo syngamy, forming a zygote that grows into a new, diploid sporophyte, and the life cycle is complete.
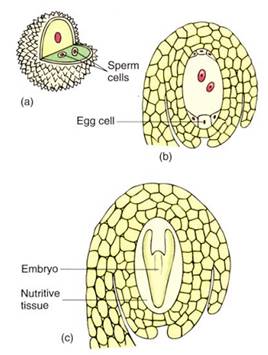
FIGURE 2 :All seed plants produce two types of gametophytes. (a) Microspores (pollen grains) develop into microgametophytes (also pollen grains). The microgametophyte body is so small that it has just thee cells and fits inside the pollen cell wall. From the outside, it is not possible to tell if a pollen grain is a microspore or a microgametophyte. The microgametophyte produces two sperm cells located within its own protoplasm. (b) The megaspore develops into a megagametophyte. It is slightly larger than a microgametophyte and has seven cells, one of which has two nuclei. One of the cells is the egg. The megaspore and megagametophyte are never released from the flower of the parent sporophyte, so they appear to be just sporophyte tissues, not one plant growing inside another. (c) After one of the sperm cells fertilizes the egg cell, the new egg cell nucleus is diploid and the cell is a zygote. It develops, by mitosis, into a new sporophyte, shown here as the embryo in an immature seed. The entire ovule develops into a seed; the ovary develops into a fruit.
Mammalian gametes are of two types: small sperm cells (microgametes) that swim, and large eggs (megagametes) that do not. This is also true of many plants and is known as oogamy. In oogamous plants, just as in oogamous mammals, sperms are produced by one type of individual and eggs by a different type of individual; hence, there are "male" or microgametophytes and "female" or megagametophytes (Fig. 2). The two types of gametophytes have grown from two types of spores: microgametophytes from micro-spores and megagametophytes from megaspores. Having two types of spores is known as heterospory. Typically just one kind of sporophyte occurs in a life cycle, and it produces both microspores and megaspores.
A life cycle like this, with two generations—sporophyte and gametophyte—is said to be an alternation of generations (Fig. 3). Because gametophytes do not resemble the sporophytes at all, this is an alternation of heteromorphic generations. This is a complex life cycle, with at least three distinct plants (one sporophyte and two gametophytes). The human life cycle, like that of all other animals, does not have anything equivalent to the haploid generation.
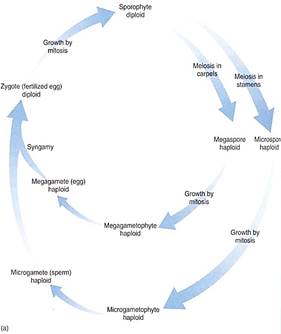
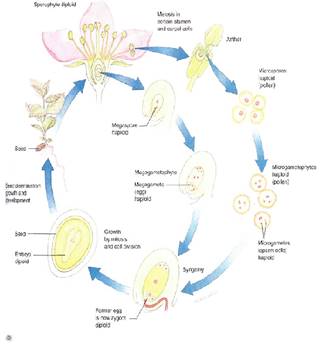
FIGURE 3 (a) :Diagram of the life cycle of a flowering plant: A sporophyte produces micro- and megaspores and each develops into micro- and megagametophytes, respectively. These produce micro- and megagametes that undergo syngamy (the sperm fertilizes the egg), producing a zygote, the fertilized egg. The zygote grows into a new sporophyte. The sporophyte generation alternates with the gametophyte generation. (b) A flowering plant life cycle showing the actual structures involved.
FLOWER STRUCTURE
A flower is basically a stem with leaf-like structures, so almost everything discussed fir vegetative shoots in Chapter 5 also applies to flowers. Flowers never become woody; secondary growth does not occur in flowers. The flower stalk is a pedicel, and the very end of the axis, where the other flower parts are attached, is the receptacle (Fig. 4). There are four types of floral appendages: sepals, petals, stamens, and carpels. Most flowers have all four types and are complete flowers.
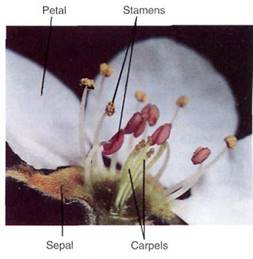
FIGURE 4:All flower parts are present and easily visible in this pear blossom. The anthers of the outermost, tallest stamens have opened, releasing their pollen. (Runk/Schoenberger from Grant Heilman).
They typically have three, four, five or more appendages of each type; for example, lilies have three sepals, three petals, three stamens, and three carpels. It is not uncommon for flowers of certain species to lack one or two of the four basic floral appendages, thus being incomplete. Flowers of willow (Fig. 5) are incomplete, lacking both sepals and petals but having either stamens or carpels; flowers of pigweed have all parts except petals.

FIGURE 9.7:In most species of flowering plants, the stamens and carpels are produced together in the same plant, but in a few species like these willows, stamens occur in flowers without carpels on one plant (a), while carpels occur in stamenless flowers on different plants (b). In contrast to mammals, the two types of plants look so much alike that they usually cannot be identified as staminate (having stamens) or carpellate (having carpels) without looking at the flowers. (a, Dwight R. Kuhn; b,]ohn D. Cunningham/Visuals Unlimited).
Sepals. Sepals are the lowermost and outermost of the four floral appendages. They are modified leaves that surround and enclose the other flower parts as they mature. Sepals are typically the thickest, toughest, and most waxy of the flower parts. They protect the flower bud as it develops, keeping bacterial and fungal spores away, maintaining a high humidity inside the bud, and deterring insect feeding (Fig. 6a and b). Sepals also protect the flower from nectar-robbing insects and birds. If flower buds develop in a protected position such as beneath a spiny cover or surrounded by regular leaves and branches, sepals are less important for protection and may be quite reduced or absent. It is not uncommon for sepals to be colorful (petalloid) and help attract pollinators. All the sepals together are fed to as the calyx.

FIGURE 6: (a) The sepals of this rose form a tight covering over the rest of the flower as it develops, protecting the inner parts. Notice that the sepals and the pedicel have glands on them, helping to deter insects. When the microspores (pollen) and megaspores (in the ovules) are ready, the sepals bend outward and the flower opens. (b) Sunflowers and daisies are actually clusters of many small flowers that together have the appearance of a single flower. In this case, the whole cluster of flowers is protected by a set of bracts that look and function like sepals. It is not necessary for the individual flowers to have protective sepals. (c) Petals, like these of the mallow, are leaf-like in being thin and flat, but in most species, they are more highly modified and less lea-flike than sepals. (d) Leaves and petals of Arbutus can be compared here; these petals are highly modified, and the five petals of each flower have fused together, forming a tubular corolla.
Petals. Above the sepals on the receptacle are petals, which together make up the corolla. Sepals and petals together constitute the flower's perianth. Petals are also "leaf-like," being broad, flat, and thin, but they differ from leaves in that they contain pigments other than chlorophyll, have fewer or no fibers, and tend to be thinner and more delicately constructed (Fig. 6c and d). Petals are important not merely in attracting pollinators, but rather the correct pollinators. Each plant species has flowers of distinctive size, shape, color, and arrangement of petals, allowing pollinators to recognize specific species. Sexual reproduction cannot occur efficiently if pollen is carried to other plants indiscriminately. Reproduction results if pollen is carried only to other flowers of the same species. If a flower has a distinctive pattern and offers a good reward such as nectar or pollen (Fig. 7), the pollinator is likely to search for and fly to other flowers with the same pattern, thereby enhancing cross-pollination; mutations are advantageous selectively if they cause flowers to have characters that are easily recognized by their pollinators. In addition to visible colors, nary flowers have pigments that absorb ultraviolet light, creating patterns only insects can.
Pollen is carried by wind in many species and by water in a few. Petals cannot attract wind or water, so loss of petals is not disadvantageous for these species. Mutations that inhibit their differentiation prevent plants from wasting carbon and energy in the construction of nonfunctional petals. Typically, petals do not develop in wind-pollinated species.

FIGURE 7:Some flowers do not produce nectar; the pollinator eats part of the stamens and pollen instead. This is more expensive metabolically for the plant because pollen is rich in protein, whereas nectar is composed of carbohydrate. In these flowers of cannonball tree (Couropita), the lower stamens are modified for edibility; they are large and showy and produce little or no pollen. The pollen-producing stamens are small and inconspicuous, easily overlooked by pollinators.

FIGURE 8: (a) A flower as seen by the human eye. (b) The same flower photographed through filters that reveal the pattern of its pigments that absorb ultraviolet light. Although we cannot see this pattern, most insects can. Distinctive patterns like this, in either ultraviolet or visible light pigments, direct the pollinator to the nectar and position the animal to pick up pollen; the markings are known as nectar guides. (Thomas Eisner).
Stamens. Above the petals are the stamens, known collectively as the androecium. Stamens are frequently referred to as the "male" part of the flower because they produce the pollen, but technically they are not male because the flower, being part of the sporophyte, produces spores, not gametes. Only gametes and gametophytes have sex.

Stamens have two parts, the filament (its stalk) and the anther, where pollen is actually produced. As part of the sporophyte, the anther is composed of diploid cells, and in each anther four long columns of tissue become distinct as the cells enlarge and prepare for meiosis . These microspore mother cells or microsporocytes continue to enlarge and then undergo meiosis, each producing four microspores (Fig. 9b; Table 1). Neighboring anther cells, in a layer called the tapetum, act as nurse cells, contributing to microspore development and maturation. Microspores initially remain together in a tetrad, but later separate, expand to a characteristic shape, and form an especially resistant wall. They are then called pollen. The anthers open (dehisce) along a line of weakness and release the pollen.
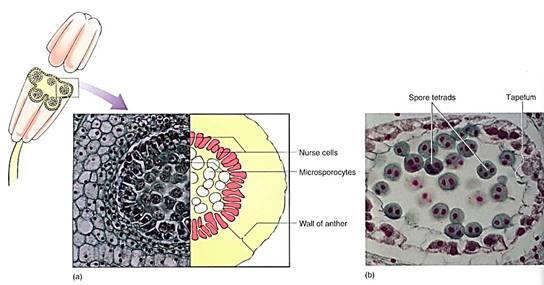
FIGURE 9: (a) Transverse section through an anther of a lily, showing the microsporocytes in prophase of the first meiotic division that will produce haploid microspores. The surrounding cells constitute the tapetum and aid in the maturation of the pollen grains (X 200). (C. I. Bernard/Earth Scenes) (b) After meiosis, the four microspores remain together temporarily, but as they develop their specialized pollen cell wall, they usually separate from each other (X 500).
The pollen wall is a cell wall; however, it is quite complex structurally. It has an inner layer called the intine, composed of cellulose, and an outer layer called the exine that consists of the polymer sporopollenin. It has one or several weak spots, germination pores, where the pollen opens after it has been carried to the stigma of another flower. Sporopollenin is remarkably waterproof and resistant to almost all chemicals; it protects the pollen grain and keeps it from drying out as it is being carried by the wind or animals. The exine can have ridges, bumps, spines, and numerous other features so characteristic that each species has its own particular pattern (Fig. 10). In many cases, it is possible to examine a single pollen grain and know exactly which species of plant produced it. Because the sporopollenin is so resistant, pollen grains and their characteristic patterns fossilize well. By examining samples of old soil, botanists can determine exactly which plants grew in an area at a particular time in the ancient past.
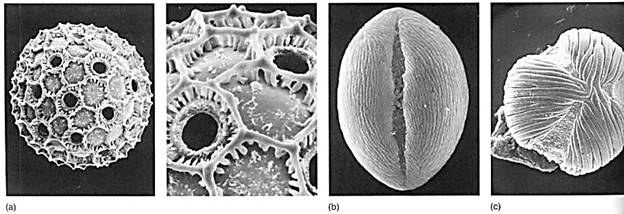
FIGURE 10 (a) Pollen grains of Cobaea, with the pollen wall forming hexagonal ridges. The numerous holes are germination pores where pollen tubes emerge after the pollen lands on a stigma (X 2000). (Courtesy of Alan Prather, University of Texas) (b) Pollen of Lycium has a single long groove from which the pollen tube emerges (X 4000). (c) Pollen of Macrolobium has three germination grooves (X 4000). (bandc, Courtesy of Beryl Simpson, University of Texas).
Carpels. Carpels constitute the gynoecium, located at the highest level on the receptacle (Fig. 11 a). Carpels have three main parts: (1) a stigma that catches pollen grains, (2) a style that elevates the stigma to a useful position, and (3) an ovary where megaspores are produced. A flower can have zero (some imperfect flowers) to many carpels; usually they are fused together into a single compound structure, frequently called a pistil. Inside the ovary are placentae (sing.: placenta), regions of tissue that bear small structures called ovules. Ovules have a short stalk, called a funiculus, that carries water and nutrients from the placenta to the ovule by means of a small vascular bundle. The ovule has a central mass of parenchyma called a nucellus. Around the nucellus are two thin sheets of cells (integuments) that cover almost the entire nucellus surface, leaving only a small hole (micropyle) at the top (11c). As in anthers, some nucellus cells, usually only one in each ovule, enlarge in preparation for meiosis; these are megaspore mother cells or megasporocytes. After meiosis, usually three of the four megaspores degenerate, and only one survives, becoming very large by absorbing the protoplasm of the other three. Megaspores differ from microspores (pollen) because the ovule and the carpel do not dehisce and the megaspore remains enclosed inside the carpel.
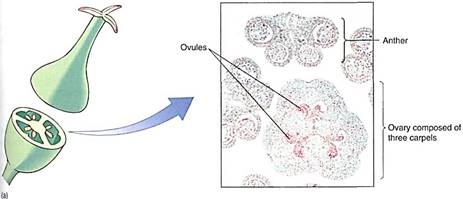
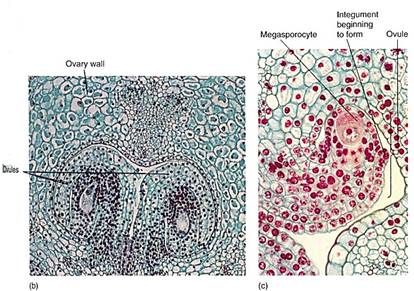
FIGURE 11 (a) In lily, the pistil consists of three carpels fused together. This lily ovary has been cut across to reveal the three ovary locules or chambers. In each chamber two ovules are visible (X 15). (b) Ovules are attached to the placenta by means of their funiculus, which contains both xylem and phloem. The central part of the ovule is the nucellus with (usually) just one megasporocyte, unlike the anthers which have numerous microsporocytes (X 50). (c) Magnification of a single ovule. Surrounding the nucellus are the integuments; here they are extremely short and do not yet cover the tip of the nucellus. Growth of the ovule has been by mitosis, but now one cell, the megasporocyte-megaspore mother cell-is preparing for meiosis (X 120). (b, M. I. Walker, Science Source/Photo Researchers., Inc.)
THE GAMETOPHYTES
Microgametophyte. Microspores develop into microgametophytes. In flowering plant species, the microgametophyte is very small and simple, consisting of at most three cells located within the original pollen cell wall (Fig. 12). The microspore nucleus migrates to the side of the pollen grain and lies next to the wall.
There it divides mitotically, producing a large vegetative cell and a small lens-shaped generative cell, which subsequently divides and forms two sperm cells. The entire microgametophyte consists of the vegetative cell and the two sperm cells (the microgametes); this is a full-fledged plant.
In about 30% of flowering plant species, the formation of sperm cells occurs even while the pollen is still located within the anther. In the majority of angiosperm species, the pollen is released from the anther at about the time the generative cell has formed, and sperm cells are not produced until after the pollen has been carried to a sigma.
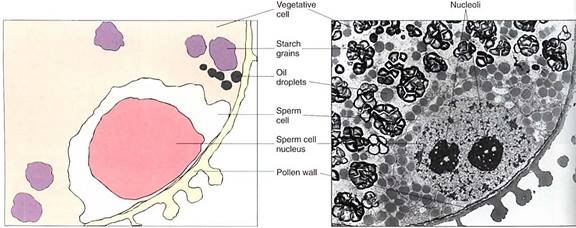
FIGURE 12 :Only with transmission electron microscopy can the details of a microgametophyte be studied well. The large cell is the vegetative cell, and the two smaller cells (only one visible here) are the sperm cells that arose by division of the generative cell. In this species, the sperm cells have no cell walls; they are separated from the vegetative cell only by plasma membranes (X 5000). (Courtesy of S. Nakamura and H. Miki- Hirosige, Kanagawa Dental College).
After the pollen lands on a stigma, it germinates by producing a pollen tube that penetrates into the loose, open tissues of the stigma (Fig. 13). The pollen tube absorbs nutrients from the stigma and grows downward through the style toward the ovary. The microgametophyte is protected and nourished by the style tissue. As the pollen tube grows downward, it carries the sperm cells to the ovule. Almost all the pollen cytoplasm is located at the tip of the pollen tubes; the rest of the tube and the pollen grain are filled with a giant vacuole. Pollen grains are too small to store much starch or protein; if they could not absorb nutrients from the style, the pollen tubes could not grow long enough to reach the ovules.
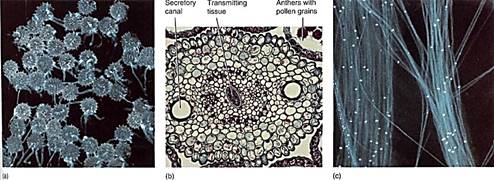
FIGURE 13 :(a) Shortly after landing on a stigma of the correct species, a pollen grain may germinate by pushing out part of the wall as a pollen tube. This penetrates the loose tissues of the stigma and style. Photographed with ultraviolet light which causes the pollen wall and callose to fluoresce and shine (X 50). (b) Tissues of the style allow the pollen tubes to grow through them easily. Some styles are hollow and lined with a rich, nutritious transmitting tissue; other styles are solid but also have transmitting tissue. Sunflower (Helianthus) style (X 80). (c) These pollen tubes have penetrated deep into the style. The bright spots are callose plugs which seal off the protoplasm at 1he tip from the empty parts of the pollen tube (X 200). (a and c, Courtesy of Allison Snow, Ohio State University).
Megagametophyte. Within the ovule the surviving megaspore develops into a megagametophyte. In one type of development, the nucleus undergoes three mitotic divisions, producing two, four, and then eight haploid nuclei (Fig. 14). The nuclei migrate through He cytoplasm, presumably pulled by microtubules, until three nuclei lie at each end and tow in the center. Walls form around the nuclei, and the large, eight-nucleate megaspore becomes a megagametophyte with seven cells, one of which is bmucleate. The seven cells are one large central cell with two polar nuclei, three small antipodal cells, and an egg apparatus consisting of two synergids and an egg (the megagamete) (Table2). Like the microgametophyte, the megagametophyte is a distinct plant. As with the pollen, the mega- gametophyte obtains all its nourishment from the parent sporophyte.
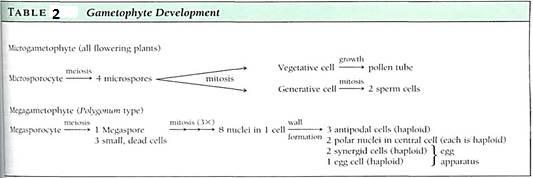
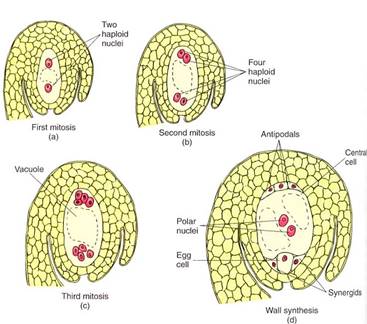
FIGURE 14 :As a megaspore develops into a megagametophyte, three sets of mitosis occur (a to c) without any cytokinesis, so the megagametophyte is temporarily an eight-nucleate coenocyte (c). Then nuclei migrate, cytoplasm accumulates around each, and walls are established (d). Several studies have shown that much of the egg is not covered by wall. The central cell is binucleate and is mostly vacuole.
FERTILIZATION
Syngamy of sperm and egg involves both plasmogamy, the fusion of the protoplasms of the gametes, and karyogamy, the fusion of the nuclei. As a pollen tube grows downward through the style toward the ovule, it is guided to the ovule's micropyle by some means. It penetrates the nucellus and reaches the egg apparatus, then enters one synergid. The pollen tube tip bursts and releases both sperm cells. One of these migrates toward the egg, and as it does so, its plasma membrane breaks down. The sperm cell loses most of its protoplasm before it reaches the egg. As the sperm fuses with the egg plasma membrane, only the sperm nucleus enters the egg. The sperm nucleus is drawn to and then fuses with the egg nucleus, establishing a diploid zygote nucleus.
Because the sperm sheds its protoplasm as it passes through the synergid, it contributes only its nucleus with the set of nuclear genes during karyogamy. The sperm does not carry mitochondria or plastids into the egg, so organellar genes from the pollen parent are rarely inherited by the zygote. Instead, the mitochondrial and plastid genes of the embryo are inherited only from the ovule parent.
In flowering plants only, the second sperm nucleus released from the pollen tube migrates from the synergid into the central cell. It undergoes karyogamy with both polar nuclei, establishing a large endosperm nucleus that is triploid, containing three full sets of genes. Because both sperm nuclei undergo fusions—one with the egg nucleus, the other with the polar nuclei—the process is called double fertilization. The endosperm nucleus is extraordinarily active and begins to divide very rapidly by mitosis, with cell cycles lasting only a few hours.
The endosperm nucleus initiates a dynamic cytoplasm and the central cell enlarges enormously, usually without cell division, into a huge cell with hundreds or thousands of nuclei (15). Finally, nuclear division stops, and dense cytoplasm gathers around the nuclei. Walls are constructed, thus forming cells. An example of this is a coconut full of "milk." The hollow center of the coconut is one single cell, and the milk is its protoplasm. The white coconut "meat" is the region where nuclei form cells. A green, immature coconut is full of milk but has almost no meat; as it ripens, the coenocytic milk is converted to a thick layer of cellular meat. All this tissue, both coenocytic and cellular, is called endosperm, and it nourishes the development of the zygote. No other megagameto-phyte forms as much endosperm as a coconut does. A more typical example is corn, in which most of the kernel is endosperm. Corn endosperm also passes through a milk stage, but most people are not familiar with it.
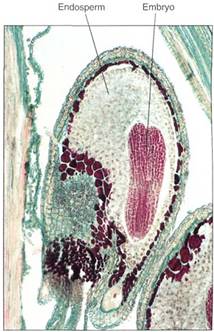
FIGURE 15: In many species, endosperm development is accompanied by some cytoplasmic division, so the endosperm is a mass of multinucleate "cells," each with variable amounts of protoplasm and nuclei and with irregular shapes. This is the only plant tissue in which nuclear and cytoplasmic divisions are so poorly coordinated, possibly because the endosperm is just a temporary tissue that is consumed by the embryo before or soon after seed germination (X 180).
EMBRYO AND SEED DEVELOPMENT
As the endosperm nucleus proliferates, the zygote also begins to grow, but always by both nuclear and cellular divisions; a coenocytic stage never occurs in the embryo. The zygote grows into a small cluster of cells, part of which later becomes the embryo proper, and the other part becomes a short stalk-like structure, the suspensor, which pushes the embryo deep into the endosperm (Fig. 16). The suspensor is usually delicate and ephemeral in flowering plants; it is crushed by the later growth of the embryo and is not easily detectable in a mature seed.
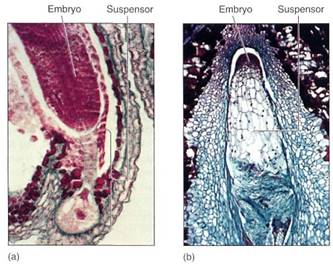
FIGURE 16 :(a) He suspensor of shepherd's purse (Capsella) has one large bulbous cell and a stalk of smaller cells. The young embryo is being pushed deep into the endosperm (X 600). (b) The suspensor of Tristerix is much more massive than that of Capsella, being larger than the embryo until the last sages of seed maturation. The embryo is still just a small ball of densely cytoplasmic cells (X 300).
The cells at one end of the suspensor continue to divide mitotically, developing into an embryo. The cells at first are arranged as a small sphere, the globular stage. The end of the embryo farther from the suspensor initiates two primordia that grow into two cotyledons indicots (see Figs. 17 and 18), such as beans and peanuts. While young, the cotyledon primordia give the embryo a heart shape; this is the heart stage. In monocots such as corn, only one cotyledon primordium grows out. Later, in the torpedo stage, the embryo is an elongate cylinder: A short axis is established, consisting of radicle (primordial root), epicotyl (primordial stem), and hypocotyl (the root/shoot junction). Finally, vascular tissue differentiates within the embryo. The epicotyl may bear a few small leaves, and the radicle often contains several primordia for lateral roots in its pericycle. Once mature, the embryo becomes quiescent and partially dehydrates, and the funiculus may break, leaving a small scar, the hilum.
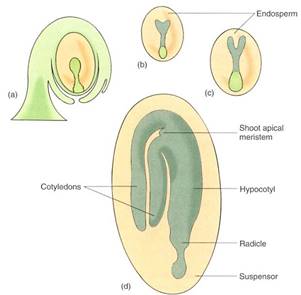
FIGURE 17 :Embryo development. (a) Globular stage. (b) Heart stage. The two mounds that cause the heart shape are the first stage of the outgrowth of cotyledons. (c) Torpedo stage. Embryo root (radicle), cotyledons, and hypocotyl are present. At this stage, the first xylem and phloem may become distinguishable in the hypocotyl. (d) Mature embryo. A shoot apical meristem is shown; in some species, even some leaf primordia and a small stem are present.
In most dicots, cotyledons store nutrients that are used during and after germination; during embryo development, the cotyledons become thick and filled with starch, oil, or protein while the endosperm, which is supplying the nutrients, shrinks. When the seed is mature, the cotyledons are large and the endosperm may be completely used up.
In monocots, the one cotyledon generally does not become thick and full; instead, the endosperm remains and is present in the mature seed. During germination, the cotyledon acts as digestive/absorptive tissue, transferring endosperm nutrients to the embryo. &me dicots are intermediate: The cotyledons store some starch and protein, but a considerable amount of endosperm remains in the seed and both methods of nutrition are used during germination. A mature seed in which endosperm is rather abundant, such as corn, is called an albuminous seed (Fig. 18). If endosperm is sparse or absent at maturity (peas, beans), the seed is exalbuminous.
The amount of embryo growth and development that occurs before dormancy sets in is extremely variable. Orchids, bromeliads, and a few other species have small, dust-like seeds in which the embryo is only a small ball of cells with no cotyledons, radicle, or vascular tissue. In seeds of most angiosperm species, all the parts are present and the epicotyl contains two or three young leaves in addition to cotyledons; these leaves can begin photo-synthesis immediately after germination (Fig. 18). In corn, the embryo is even more advanced: It contains as many as six young leaves while in the seed. A fully mature corn plant often has only 10 or 12 leaves. Over half of the leaf production of the new sporophyte occurs while it is embedded in the parental gametophyte, which is itself embedded in the previous sporophyte.
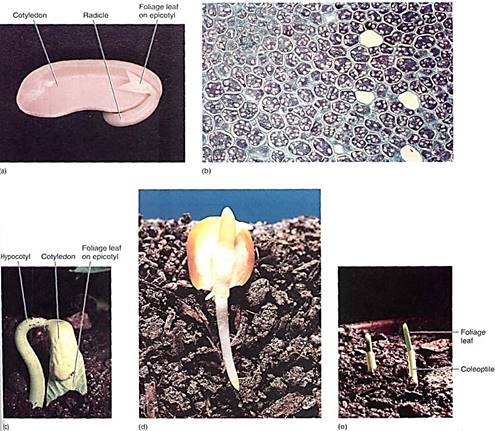
FIGURE 18 :(a) This bean seed has begun germinating; the radicle has extended and will develop into a taproot. The two cotyledons (one was removed) were digesting their carbohydrates and proteins and transporting nutrients to the root and shoot apical meristems. Beans have a well-developed Itpicotyl with several partially expanded leaves; one is visible here. (b) These cells of bean cotyledon are full of starch (stained pink) and reserve protein (stained blue) (X 100). (c) The small leaves present on the epicotyl are expanding during germination; because they were rather completely formed before the embryo became dormant, the leaves can now mature rapidly and photosynthesis begins almost as soon as germination is complete. Notice how the hypocotyl is curved This allows the leaves to be dragged gently up through the soil, protected by the cotyledons, (d) Most of a corn seed is the endosperm; the embryo is less than half the volume of the seed. The one cotyledon is large and contacts the endosperm. During germination it secretes digestive enzymes into the endosperm and absorbs the resulting monomers. (Barry L. Runk from Grant Heilman) (e) Corn seeds also produce many small leaves before becoming dormant, and fey can begin photosynthesis immediately after germination. The leaves are protected from the soil by a tubular sheath called a coleoptile.
The embryo and endosperm develop from the zygote and megagametophyte central cell, respectively, both located in the nucellus of the ovule. Soon after fertilization, or even before, synergids and antipodals break down in most species. The nucellus expands somewhat but later is crushed by the expansion of the embryo and endosperm and is usually not detectable in mature seeds.
The integuments that surround the nucellus expand and mature into the seed coat (also called the testa) as the rest of the ovule grows. In their last stages of maturation, they may become quite sclerenchymatous and tough (Fig. 19).
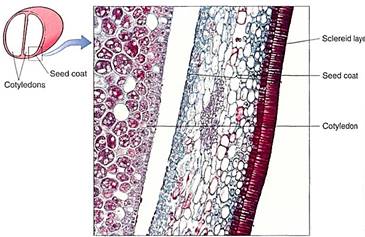
FIGURE 19:This seed coat of bean has an outer layer of sclereids and inner parenchymatous layers. The cotyledon is part of the embryo, and you might expect to see endosperm or nucellus between the cotyledon and seed coat, but those have been crushed during the later stages of seed development (X 200).
FRUIT DEVELOPMENT
As the ovule develops into a seed, the ovary matures into a fruit. Development varies with the nature of the carpels as well as the nature of the mature fruit. The stigma and style usually wither away, as do sepals, petals, and stamens, although they may persist at least temporarily (Fig. 20). Often three layers become distinct during growth: The exocarp is the outer layer—the skin or peel; the middle layer is the mesocarp, or flesh; and the innermost layer, endocarp, may be tough like the stones or pit of a cherry or it may be thin (Fig. 21). The relative thickness and fleshiness of these layers vary with fruit type, and often one or two layers are absent. The entire fruit wall, whether composed of one, two, or all three layers, is the pericarp.

Figure 20: In apples, the petals die and fall off after pollination. Stamens and sepals die and dry out, but they remain present while carpels and receptacle develop into a fruit. In most species, sepals, petals, and stamens all abscise after pollination.

FIGURE 21 :Peach fruits have all three fruit layers: Their endocarp is a solid mass of sclerenchyma around the seed, the exocarp is the skin, and the mesocarp is the flesh. Fruits with endocarps like this—a stone or pit—are drupes. (Jonas Kahn).



|
|
|
|
اكتشاف تأثير صحي مزدوج لتلوث الهواء على البالغين في منتصف العمر
|
|
|
|
|
|
|
زهور برية شائعة لتر ميم الأعصاب التالفة
|
|
|
|
|
|
العتبة العباسية تطلق مسابقة فن التصوير الفوتوغرافي الثانية للهواة ضمن فعاليات أسبوع الإمامة الدولي
|
|
|
|
لجنة البرامج المركزيّة تختتم فعاليّات الأسبوع الرابع من البرنامج المركزي لمنتسبي العتبة العباسيّة
|
|
|
|
قسم المعارف: عمل مستمر في تحقيق مجموعة من المخطوطات ستسهم بإثراء المكتبة الدينية
|
|
|
|
متحف الكفيل يشارك في المؤتمر الدولي الثالث لكلية الآثار بجامعة الكوفة
|