
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Plants in Relation to Their Habitats |


|
|
Read More
Date: 23-10-2016
Date: 30-10-2016
Date: 20-10-2016
|
Plants in Relation to Their Habitats
The habitat is the set of conditions in which an organism completes its life cycle. For migratory animals, the winter area, summer area, and migration routes are all habitat components. No plant is migratory, but portions of plants are: spores, pollen, fruits, seeds, and vegetative propagules.
How much of the surroundings should be considered part of the habitat is debated. Many factors do not appear to affect certain plants at all. The presence of small herbs on the forest floor does not seem to influence the large trees; their presence or absence has little effect on the mineral nutrition of the trees or on their pollination (Fig. 1). An experiment might remove all the small annuals and then examine whether the trees are affected, but it would be difficult to measure the growth of whole trees, especially their roots, and it might take years to see any effect on the trees' metabolism. The small herbs may be important; perhaps they harbor spiders that catch insects that would otherwise kill the trees' seedlings. If so, then removing the herbs might cause an increase in the insect population and decreased survival of tree seedlings; with fewer seedlings of this species, it might be possible for the seedlings of a different species to survive better. After many years, the forest composition would be changed as a result of the removal of the herbs.

FIGURE 1: The redwoods here completely dominate the habitat, each tree containing many times the bulk and volume of any single herb. They provide filtered light and protection from wind for the understory plants, which are incapable of tolerating full sunlight. But do the herbs affect the trees in any way? Yes: The redwood seeds germinate only after a low, cool fire fueled by the understory herbs and shrubs
On the other hand, we do know that many components impact others directly. Pollinators are critically important aspects of the habitat for the plant species they pollinate, and any disease organisms or predators that prey on those pollinators are also important to the plant. Those aspects of the habitat that definitely affect a plant constitute its operational habitat, whereas all the components whether with known effect or not, are its habitat.
For example, consider the redwood forests in California. It was discovered that there were no redwood seedlings in the national parks, and redwood seeds were not germinating. After a recent forest fire, redwood seeds sprouted and grew vigorously. The fire not only stimulated the seeds to germinate but, by burning the understory plants, released minerals that increased the soil fertility. We now realize that the shrubs and herbs are vital to the success of the giant redwoods: They fuel quick, cool fires that are too low to damage the large redwoods but are necessary for seed germination. The policy of preventing forest fires—a natural factor of this ecosystem—was harming the redwoods.
Habitat components are of two types, abiotic and biotic. The abiotic components are nonliving and are physical phenomena: climate, soil, latitude, altitude, and disturbances like fires, floods, and avalanches. Biotic components are the living factors: the plant itself, other plant species, and species of animals, fungi, protests, and prokaryotes.
ABIOTIC COMPONENTS OF THE HABITAT
Climate. Climate is critically important to all organisms; most species are restricted to certain regions primarily because they cannot live in climatic conditions outside these regions .Climate itself has many components—temperature, rainfall, relative humidity, and winds being just a few.
The average temperature of a habitat is not as important as its extremes: the lowest winter temperature and the highest summer temperature. Many species of bromeliads, aroids, and orchids are restricted to the tropics because those are frost-free habitats. Rainforests along the west coast of California, Oregon, and Washington receive adequate rain, but the freezing winters prevent them from being suitable habitats for most tropical species. On the other hand, many temperate trees must have a winter dormancy period accompanied by weeks of subfreezing temperature in order to be vernalized and bloom. If cultivated in areas with warm winters, the plants grow and survive well but do not reproduce. The highest or lowest temperatures during a plant's lifetime are important. If a tree species must be ten years old before it can flower but killing temperatures occur every eight or nine years, the species cannot survive there the growing season of an area is often determined by the date of the last severe, killing frost in the spring and the first killing frost of autumn. The length of the growing season must be adequate for sufficient photosynthesis, growth, development, and reproduction; if not, a reproductive population cannot survive even if it can tolerate the temperature extremes of winter and summer.
Moisture occurs as rain or snow or as hail that supplies water but also damages leaves, buds, flowers, and animals. Habitats range from extremely dry (deserts) through progressively more moist all the way to marshes, lakes, and rivers that are virtually all water. Just as with temperature, the total amount of precipitation may not be as important as the seasonal extremes or the timing of the precipitation. A constant drizzle that occurs almost year- round supports certain types of ecosystems, whereas the same amount of rain, distributed as just winter snowfall and occasional summer thunderstorms, results in a different type of ecosystem.
Numerous metabolic processes respond proportionally to abiotic factors. Once there is sufficient moisture for marginal survival, increased amounts of water produce increased growth and reproduction. There is usually an upper limit: With too much water, roots drown for lack of soil oxygen (Fig. 2a). Between the low and high extremes is the tolerance range of the organism. Ranges vary greatly from species to species (Fig. 2b). Some are extremely broad: Most temperate plants across the northern United States and Canada, especially in the Midwest, tolerate summer highs of over 100°F and winter lows below —40°F. Plants of southern Florida, Puerto Rico, and Hawaii tend to have much narrower tolerance ranges, being killed both by cool temperatures and by hot ones.
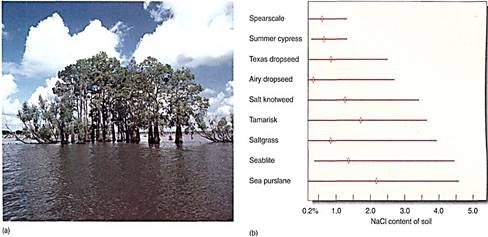
FIGURE 2: (a) Although most plants would grow better if given more water, the swampy bayous of southern Louisiana contain so much water that most plants cannot grow; their roots drown. Bald cypress (Taxodium) is well-adapted, however; its roots form "knees" filled with aerenchyma that allow atmospheric oxygen to diffuse into the submerged roots. (b) Plants vary in their ability to tolerate salty soil. These species occur around salt flats in Oklahoma and Kansas; sea purslane and seablite can survive even in the presence of strong salt concentrations, whereas others tolerate less. By being tolerant of soil salt, these plants can occupy habitats on the edges of salt flats which are free of other plants. Not shown are the many species that have virtually no salt tolerance. Diamond marks indicate the optimal concentrations.
Soil Factors. Soils are formed by the breakdown of rock. Initially the resulting soil is thin and virtually identical to the parent rock in its chemical composition; consequently, young soils are variable in the amounts of macronutrients and micronutrients they have available. Because nitrogen is not a significant component of any type of rock, all young soils are deficient in it.
The first plants that invade a new soil, called pioneers, must be able to tolerate severe conditions. The soil is sandy, with relatively large particle size, and most minerals are still locked in the rock matrix. The soil has little water-holding capacity, and the first plants have no neighbors to help moderate the wind, provide transpired humidity, or otherwise temper the environment (Fig. 3).

FIGURE 3: This "soil" consists mostly of rather large rock fragments with little water-holding capacity and very few dissolved essential elements. The plants growing here, pioneers, not only tolerate these conditions but actually change them. The acids they release as they decompose greatly accelerate chemical weathering and soil formation.
Pioneer plants often are associated with nitrogen-fixing prokaryotes; many lichens contain cyanobacteria, and angiosperms have root nodules. As the pioneers live on the soil, they change it significantly; carbon dioxide from root respiration produces carbonic acid and accelerates chemical weathering. Dead plant parts such as leaves, fruits, roots, and bark become substrates for soil organisms, and their decay contributes humus, greatly increasing the soil's water-holding capacity. Roots may penetrate to the bedrock, entering larger cracks and then expanding and breaking the rock physically.
After many years, a thick soil may result that has a distinct soil profile with three layers or horizons (Fig. 4). The A horizon is uppermost and is sometimes called the zone of leaching; it consists of litter and debris, and as this breaks down, rainwater washes nutrients from it downward into the next layer, the B horizon, or zone of deposition. The B horizon is the area where materials from the A horizon accumulate. It is rich in nutrients and contains both humus and clay. Below is the C horizon, composed mostly of parent rock and rock fragments.
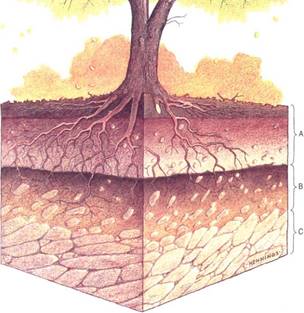
FIGURE 4: Most soils show three horizontal layers, or horizons. The relative thicknesses of the A horizon (zone of leaching) and B horizon (zone of deposition) depend on many factors, the abundance of vegetation and humus being especially important.
Whereas young soils differ because of the chemical nature of their parental rock, older, more mature soils are less diverse. As the rock weathers, essential elements are absorbed by roots and become trapped in the plant body; nonessential elements are leached away. As the plants or their parts die, they fall to the ground and decay slowly, releasing the essential elements which re-enter the soil. There they are taken up again. As a result, essential elements cycle repeatedly, alternating between organisms and the A and B horizons, whereas other elements are gradually washed downward into the water table and removed by underground water flow.
Latitude and Altitude. Latitude contributes many factors to the abiotic environment. At the equator all days are 12 hours long, no seasonal variation occurs, and plants cannot measure season by photoperiod. At progressively higher latitudes to either the north or south, summer days become progressively longer, as do winter nights (Fig. 5; Table 1). Above the Arctic and Antarctic Circles, summer days are 24 hours long, as are winter nights. At intermediate and higher latitudes, day length is an excellent indicator of season, and some species are sensitive to photoperiod.
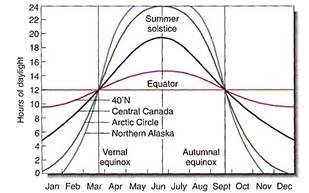
FIGURE 5: The line for day length at 40 degrees north, which corresponds to the center of the United States, shows that the longest day (summer solstice, about June 21) is just less than 15 hours long, whereas the shortest day (winter solstice, about December 21) is only about 10 hours long. Closer to the equator, the difference between the longest and shortest days is less, and near the equator tie difference is too little for plants and animals to be able to use it as a seasonal indicator.
The amount of light energy that strikes a given area of Earth's surface each year also varies with latitude; in equatorial regions, the sun is always nearly overhead, and each square meter of surface receives a maximum amount of radiation (Fig. 6). At higher latitudes the sun is only rarely overhead, usually only near midsummer. At other times, when the sun is low, light strikes the Earth obliquely and less energy is received per square meter; even at noon on winter days the sun is low in the sky and the days are not bright. At high latitudes, temperatures fluctuate greatly on both a daily and a seasonal basis. Soil formation is slow in the cold latitudes, and often what soil has formed is blown away by strong winds. In much of Alaska and Canada, soil was scraped away during the ice ages 18,000 years ago.
Regions of high altitudes on mountain tops are similar to those at high latitude. There are high winds and poor soil; much or all of the year is cold, and the growing season is short. Water may be present mostly as snow and ice, so physiological drought is frequent the areas may have varying day lengths, depending on their latitude. An additional stress present in high-altitude habitats is intense ultraviolet light. High altitudes are above much of Earth's atmosphere and thus are not fully shielded by ozone, oxygen, carbon dioxide, and water vapor.
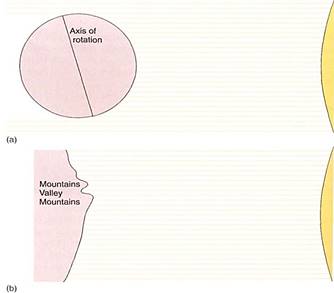
FIGURE 6: (a) During winter in the Northern Hemisphere, the North Pole is pointed away from the sun and the amount of energy received per square meter is at a minimum. The South Pole points toward the sun and the Southern Hemisphere receives more direct light, resulting in a large amount of energy per square meter. Six months later the positions are reversed. (b) The variability of energy received per square meter of surface is affected by topography. Away from the equator, the side of a mountain or valley that faces the equator also faces the sun and receives more energy. The sides that face the poles are cooler. Very often, the two sides of a valley running east to west have different vegetation.
Disturbance. Disturbances are phenomena such as fires, landslides, snow avalanches, and floods; they produce a significant, often radical change in an ecosystem quickly (Fig. 7). Disturbances affect the biotic factors directly, often completely eliminating many or all individuals from an area and also altering the soil, but they have little or no impact on other abiotic factors such as climate, latitude, or altitude. The elimination of large numbers of individuals by a disturbance alters species relationships in the ecosystem. Man-made disturbances have been caused by insecticides, herbicides, hunting, and habitat destruction (Fig. 8).

FIGURE 7: (a) Fires have a significant impact on the ecosystems. They kill not only plants but also pollinators, herbivores, and pathogenic fungi and bacteria. Fire also releases minerals back to the soil and decreases shading. A quick, cool fire does not damage rhizomes, tubers, seeds, or trees with thick bark. (SJ. Krasemann/Peter Arnold) (b) within a few months after a fire, mosses, grasses, and wildfowls have begun regrowth. (c and d) The eruption of Mt. St. Helens radically altered the surrounding ecosystems, but there too plants, animals, and other organisms are recovering. (c, M. Shafer/Peter Arnold; d, Larry Nielsen/Peter Arnold)
Fire is a natural, common component of many dry ecosystems. With little moisture, fallen leaves and twigs that a thick layer of debris builds up. The living plants tend to have waxy cuticles and water-proofing resins that make them especially flammable. Lightning storms often occur without rainfall, starting fires that burn rapidly and cause great destruction. Many of the plants and animals of such ecosystems have become fire-resistant as a result of natural selection caused by frequent fires. The bark of certain species of pine trees is so thick that a rapidly moving, moderate fire does no damage to the vascular cambium and other living tissues; the lower portions of the trunk have no branches because of self-pruning, so flames cannot reach high enough to ignite needles; only herbs and small shrubs are burned. Furthermore, the cones of lodgepole pine and jack pine open only after being exposed to the heat of fire; this adaptation results in the release of seeds after many competing plants and predatory animals have been killed and the forest floor is open and sunny.

FIGURE 8: Humans destroy habitats on a massive scale; once this hydroelectric dam is completed, it will permanently flood the valley upstream and convert a tropical forest to a lake. Downstream, the flooding that had been a natural part of the habitat, important in maintaining the community, will be eliminated. The availability of electrical power will almost certainly result in an immigration of people, causing further land clearing and habitat destruction.
Also the soil is enriched by the minerals in the ash, making it an ideal site for pine seedlings. However, if fires do not occur frequently enough, usually because foresters put them out, understory shrubs and small trees grow tall and large; so much brush and dead wood accumulate that when fire does occur, it is extremely hot. As it burns to the top of the shrubs and understory trees, flames may reach the lowest branches of the pines, igniting the crowns (Fig. 9). Once this occurs, the fire can spread rapidly through the canopy of trees, killing them.
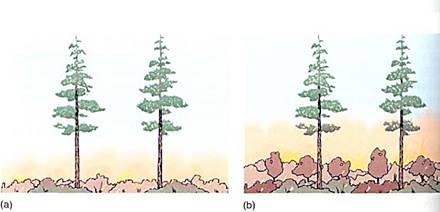
FIGURE 9: (a) If fire occurs frequently, understory shrubs are burned back before they become tall and there is always a large space between them and the lowest branches of the dominant trees. Fire cannot get high enough to ignite the tree canopy. (b) If fire occurs infrequently, understory shrubs become tall, reaching the lowest branches of the dominant trees. Fire can then burn upward from the shrubs into the canopy, igniting the highly flammable needles, twigs, and cones. Even if the trees are not killed outright, their shoot meristems are destroyed and no further growth is possible.
Many grasses have adaptations that permit them to benefit from fires. Many prairie grasses in the Midwest and the saw grasses of the Florida everglades grow in dense dumps with their shoot tips and leaf primordia at or below ground level, protected from fire by soil and the living bases of leaves. Leaf tips may be dead and dry, and when fire occurs, the dead portions burn, releasing their minerals, but the bases of the plants are unharmed.
Annuals and short-lived plants do not survive fires, but their seeds, buried underground, do. Plants with bulbs, rhizomes, tubers, or corms easily survive small fires.
BIOTIC COMPONENTS OF THE HABITAT
The Plant Itself. An individual itself, just by being in a habitat, modifies the habitat and is a part of it. Habitat modification may be beneficial, detrimental, or neutral to the continued success of that species in the habitat. In the beech/oak forests of the northern United States, the trees modify the habitat by producing a dense canopy that results in a heavily shaded forest floor (Fig.10 a). With such low light levels, few seedlings grow well, but two that do are those of oaks and beeches. As a result, mature trees create a habitat that suits their seedlings and aids their own successful reproduction.

FIGURE 10: (a) In this forest, the two dominant species, beeches and oaks, alter the habitat so that it is suitable for their own seedlings. Their seeds can germinate and grow in the heavy shade they provide, but the seedlings of many other species cannot survive in such low light. (Grant Heilman Photography) (b) Although much more open and sunny than a beech/oak forest, a pine forest is still too shaded for pine seedlings. Pines seem to alter the habitat adversely for their own long-term survival, but the needles they drop are highly flammable and are the main cause of frequent fires that kill oak seedlings but not pine seedlings. (William E. Ferguson) . (c) For several years, a pine seedling remains short with the shoot apical meristem well protected at ground level, surrounded by moist, nonflammable living leaves. It can survive quick, cool fires and builds an extensive root system. After about 3 to 7 years, the seedling begins to grow upward very rapidly, with most energy going into stem elongation and little to the roots. Within just 2 or 3 years, the tree may be more than 18 feet tall and the shoot apical meristem is out of danger from fires.)
Pine forests are more open, but still the forest floor is shaded by mature pines. Pine seedlings, however, need full sunlight and do not grow well below the canopy of older pines. Seedlings of other species flourish in these conditions and crowd out the few pine seedlings that may occur. Pine trees modify their habitat adversely for their continued success; only disturbances can create the open habitats needed for pine seedlings (Fig. 26.12b and c).
As glaciers retreat, they leave behind moraines—great mounds of rubble, sand, and boulders. The soil is poor, with no humus and few available nutrients, but pioneer species such as alder; Dryas (in the rose family), willow, and fireweed are able to colonize recently exposed moraines (Fig. 11). They survive in the open conditions and tolerate low levels of nutrients; the alders and Dryas have root nodules containing symbiotic nitrogen-fixing bacteria that supply nitrogen. Within a few years, the decay of their leaves and bark has enriched the soil sufficiently that sitka spruce and western hemlock become established. After this, however, the spruce/hemlock forest creates too much shade and eliminates the pioneers that have altered the habitat to their own detriment by enriching it.

FIGURE 11: As a glacier retreats, the rubble and sand left behind is extremely poor soil, and of course the climate is usually harsh. However, several species of pioneers can grow here, and their activity enriches the soil, permitting invasion by less hardy species. Mt. Rainier National Park, Washington.
Other Plant Species. When several individuals, of either just one or several species, occur together, the possibility for interaction is created. If the interaction is basically beneficial £r both organisms, it is described as mutualism, but if it is disadvantageous, it is competition. Competition is a situation in which two populations do not grow as well together as they do separately because they use the same limited supply of resources. Many plants are believed to compete with others for light, soil nutrients, water, and the attention of pollinators and seed dispersers, among other things. If a single plant were allowed to grow by itself, in many cases it might grow more rapidly, become larger, and produce many more gametes than it would if other plants were nearby. Roots of the other plants might grow among its roots and remove water and nutrients. The competitors might grow taller than it does and then intercept sunlight by putting their leaves above its leaves. Their flowers, even if they did not produce more nectar, might still distract its pollinators such that its pollen would be carried to the stigmas of the wrong species and its own stigmas would receive foreign pollen.
The role of competition has been extensively debated. One theory postulates that the result of competition is competitive exclusion; whichever species is less adapted is excluded from the ecosystem by superior competitors. The species that get sunlight and other resources win; those that do not, lose and are eliminated. If this is true, then very little competition occurs in a typical ecosystem; each species is assumed to be adapted to a particular set of conditions, a niche, that no other species is adapted to use as efficiently, For example, some species are adapted to full sunlight, others to partial shade (Fig. 12). In an ecosystem, the former must be a canopy tree and the latter an understory species This theory of little competition predicts that if certain species are removed, the others do not benefit from the unused resources because they are not adapted for them. This is sometimes found to be the case.
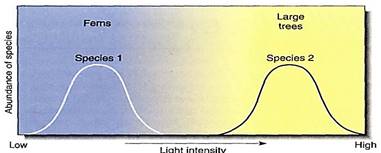
FIGURE 12: Both ferns and large trees need light, but many fern species cannot tolerate high light intensity and must have partial shading. Large trees often must have intense light; they do poorly if shaded. Growing together, they do not compete for sunlight; each uses a portion of the resource the other does not use. If either were removed, the other would not grow better
The concept of niche is difficult to define exactly; basically it refers to the set of aspects of the habitat that directly affect a species. For example, a particular marsh species occupies a particular semiaquatic niche defined by a range of soil moisture or flooding, a range of seasonal rainfall, a range of temperature, a paucity of root grazing due to lack of swimming herbivores, and the presence of appropriate pollinators. Another species of marsh plant may grow in the same general geographical area, even the same marsh, but occupy a different niche because it may have a different pollinator or may grow in areas of the marsh that are slightly more acidic than the microhabitats occupied by the first species. As long as even one factor differs, they occupy different niches.
As an alternative to the theory of competitive exclusion, a second theory postulates that species overlap in their tolerance ranges (Fig. 13a), and, when grown together, each has exclusive use of the portion of the range not used by the other. In the overlap zone where the habitat is suitable for both species, usage is determined by competition: The one that is more adapted occupies the overlap zone exclusively. The weaker of the two species occupies only part of its potential niche. The two can coexist, but if either is removed, the other then has its full range of resources available and grows better. Examples of this type of competition have been found to occur.
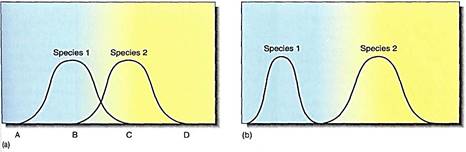
FIGURE 13: (a) Two species compete if they can use the same resource. Here, species 1 would have exclusive us of the resource under conditions A to B, whereas species 2 would have exclusive use between C and D. With conditions between B and C, they would compete. If the resource were water availability, species 1 might grow in drier areas and species 2 in wet areas, and both could grow in moist areas. Whether they both actually live in moist areas depends on how well they compete; ere may be so efficient as to exclude the other. In areas of competition, one survives better if the other is removed. (b) Over time, mutations that improve the ability of a species to compete may be selected. If such mutations do not occur or if the competitor is too efficient, then mutations may be selected that restrict the efficiency of the species in areas where it does well. In this case, species 1 evolves such that it is specialized in dry habitats and now cannot live in moist conditions. This may increase its total fitness, because the capacity to live in moist environments involves more than just the ability to use abundant water; moist environments also have more insects and fungi that require expensive defense mechanisms.
A long-term result of competition should be species modification by natural selection. If two species compete with partial tolerance-range overlap, then mutations are beneficial if they allow each to use more efficiently its exclusive portion of the range. Through time, the species diverge until competition is reduced (Fig. 14b).
The geographic ranges of most populations are extensive enough that they contain a diversity of biotic and abiotic factors: Each may include hills, valleys, and plains; rocky soil and rich soil; grasslands and open woodlands. Because of this diversity, a population may be competing for sunlight in one part of its geographic range but competing for water in another and lor other factors in still another. The ecosystem diversity causes different subpopulations to specialize for certain features, especially if gene flow is not rapid and thorough. This could be the beginning of divergent speciation, of course, but long before the various subpopulations could be called subspecies, they would be considered ecotypes; each specialized in response to particular ecosystem factors at its locality. The various ecotypes resemble each other so strongly that they clearly belong to the same species but have enough differences for ecologists and taxonomists to suspect that separate ecotypes exist. Experiments are necessary to prove that natural selection is in fact altering the genome, because phenotypic differences can result simply from the growing conditions themselves. For example, beans grown in rich soil with adequate water are strikingly different from those grown in poor soil with little water and other stresses. To test if ecotypes really exist, transplant experiments are performed: Plants from each site are transplanted to the alternate site and plants from both sites may be grown together in a common garden at an intermediate site. If the transplanted individuals take on the phenotype of the naturally occurring plants at that site, then genetic factors were not important, just stresses. But if the transplanted individuals retain their original phenotype or if they die, then genetic divergence had begun.
Also, success in competition depends on factors other than the ones involved in the competition. Species A may grow rapidly and shade species B when water is abundant, but species B may be more drought-tolerant and can outgrow and outcompete species A when water is scarce. Water availability may vary over the geographic range such that A dominates in wet areas and B in dry areas. Also, because rainfall varies from year to year, species A may be more common some years and species B more abundant in other years.
Animal ecology includes a variety of negative interactions in addition to competition; predation—the killing of prey—and parasitism are two examples. Insectivorous plants do trap and kill insects, deriving nitrogenous compounds from them, but no plant kills and digests other plants in a predator-prey relationship. Parasitic plants are not uncommon. Mistletoes are familiar examples, and hundreds of species of parasitic plants in at least 12 families are known.
Numerous animals, fungi, protists, and bacteria are saprotrophs, living on dead organisms; vultures, maggots, wood-rotting fungi, and decay bacteria are examples. No plant species of any type is known to attack and digest dead plant or animal material; roots do absorb the minerals, nitrates, and sulfates that are released from decomposing humus, but the roots do not participate in the enzymatic attack.
Organisms Other Than Plants. Animals, fungi, and bacteria are obviously important biotic aspects of a plant's habitat. Interrelationships between plants and these other organisms can be either beneficial or detrimental for one or both partners.
Plants and animals have many relationships that are examples of mutualism, in which both species benefit. Most instances of pollination are mutualistic—the animal receives nectar or a portion of the pollen and the plant benefits from pollen transfer. Seed dispersal by fruit-eating animals (frugivores) also benefits both species so long as the animal does not chew the seeds and digest the embryos.
A famous example of mutualism is the association of certain ants and acacias in tropical Central America. Acacias are small trees that have enlarged, hollow thorns at their leaf base. The thorns are used as ready-made, stout, waterproof homes by ants (Fig. 14). Because of the large number of thorns on a single plant, the ants benefit from having abundant housing, but in addition, acacias produce nectar and their leaflet tips are modified into golden yellow food bodies (called Beltian bodies) that are filled with glycogen. The ants receive both housing and food. The plant benefits because this species of ant is aggressive; they patrol the plant and attack any animal that touches it, even large mammals. If the leaves of another plant come close to an Acacia, the ants destroy the leaf, keeping the Acacia unshaded. As a result, the plant is free of pathogenic fungi and insects and it glows in full sunlight. This mutualism is obligate; the plant cannot survive in nature without the ants, and the ants are not found away from the plant. A similar mutualism occurs between ants and Hydnophytum in Southeast Asia (Fig. 15).
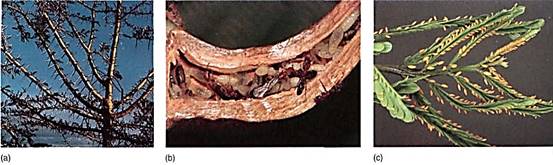

FIGURE 14: (a) Ant Acacia plant. (b) These thorns on an ant Acacia are swollen and hollow and serve as excellent nests for ants. They form as part of the Acacia's own normal development; the presence of ants does not induce their formation. (c) The leaftips of ant acacias develop as Beltian bodies (yellow), food bodies rich in glycogen. (d) As long as an acacia is occupied by ants, all other vegetation is kept cleared away, but if the ants are killed with insecticide, the acacias are overgrown quickly. (a, Zig Leszczynski/Earth Scenes; b, Robert and Linda Mitchell; d, Pau feeny)

FIGURE 15: The shoot base of Hydnophytum swells and forms hollow chambers that become occupied by ants. There are two types of chambers: Those with smooth walls are used as living quarters, and those with warty walls are used as waste dumps by the ants. The warts on the walls are absorptive cells; the walls absorb nitrogenous compounds as the wastes decompose.
Commensal relationships, in which one species benefits and the other is unaffected, are also common between plants and animals. When birds build nests in trees, the birds benefit and the tree is (usually) unharmed. When sticky fruits or seeds, such as cocklebur, stick to an animal's fur or feathers and then are dispersed, the plant benefits and the animal is unharmed. One-sided negative relationships also occur: Animals trample and kill small plants without being affected. Competition, in which both parties are adversely affected, probably is not common between plants and animals.
Predation is a relationship in which one species benefits and the other is harmed; the species that benefits seeks out the other and uses it specifically for food or some other form of resource. Animals that eat plants are herbivores and the process is hertivory, but it is often more precisely delimited as browsing (eating twigs and leaves of shrubs—deer, giraffes) or grazing (eating herbs—sheep, cattle). Insects also lay eggs in plants, then their larvae feed on plant tissues as they tunnel through them. Aphids and spittlebugs suck sap rather than chew tissues. Insects and birds both harvest leaves and twigs for nest construction.
Many interrelationships between plants and fungi or bacteria are harmful to the plant, but the fungi and bacteria are described as being pathogenic rather than predatory. Either may cause mild disease or be so virulent that they kill the plants quickly. A large percentage of the microbes are saprotrophs, living on dead plant tissues such as leaves, logs, fallen fruit, and sloughed bark. This benefits the fungi and bacteria without harming the living plants, and it can actually help them. This process speeds up the release of mineral nutrients, especially nitrogen compounds, enriching the soil. A case of plants attacking and parasitizing fungi may be known. Orchid seeds are tiny and lack chlorophyll; they remain moribund until invaded by soil fungi, then they turn green and grow well. It had been assumed that the seedling received nutrients and perhaps growth factors from the fungus and in return provided it with carbohydrate. However, tests have not revealed any benefit to the fungus, just to the plant.
Examples of plant-fungus mutualism are well known and recently have been shown to be of much greater importance than ever before suspected, as in the case of mycorrhizae. The mycorrhizal fungus transports phosphate into the plant and receives carbohydrate; both benefit. Many plant species grow only poorly in nature if the soil fungi are killed with a fungicide, and it has been postulated that one reason the plains of the American Midwest lack trees is because the type of mycorrhizae beneficial to trees cannot compete well with the mycorrhizae beneficial to grasses. A complex relationship has been discovered with Indian pipe (an achlorophyllous parasitic plant) and a mycorrhizal fungus: Indian pipe parasitizes the fungus, drawing nutrients from it, and the fungus in turn obtains carbohydrates from its other mycorrhizal partners that are chlorophyllous and photosynthetic. Indian pipe basically parasitizes other plants, using the mycorrhiza as a bridge.
The non-plant organisms add a great deal of complexity to a plant's habitat, and numerous types of interrelationships are possible. Only those relationships involving plants have been mentioned here, but the animals are competing with each other, as are the fungi and bacteria. Also, interactions occur between animals, fungi, protists, and bacteria. The ecosystem is extremely complex, and it is virtually impossible to predict how the disruption of one part might affect other parts. Although the operational habitat may be simple, the real habitat contains so many factors linked to so many other factors that we must be careful in our treatment of ecosystems.



|
|
|
|
حمية العقل.. نظام صحي لإطالة شباب دماغك
|
|
|
|
|
|
|
إيرباص تكشف عن نموذج تجريبي من نصف طائرة ونصف هليكوبتر
|
|
|
|
|
|
قسم الشؤون الفكرية يباشر باستعداداته لإقامة الدورة القرآنية لطلبة العلوم الدينية الأفارقة
|
|
|
|
بمشاركة قرّاء من البصرة... المَجمَع العلميّ يُقيم محفلَ عرش التّلاوة في صحن مرقد أبي الفضل العبّاس (عليه السّلام)
|
|
|
|
قسم الشؤون الفكريّة يُقيم جلسةً حواريّة لممثلي المواكب في ذي قار
|
|
|
|
قسم الشؤون الفكرية يعد حزمة من الدورات الدينية والثقافية المتنوعة
|















