























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Pseudomonas
 المؤلف:
SILVA, N.D .; TANIWAKI, M.H. ; JUNQUEIRA, V.C.A.; SILVEIRA, N.F.A. , NASCIMENTO , M.D.D. and GOMES ,R.A.R
المؤلف:
SILVA, N.D .; TANIWAKI, M.H. ; JUNQUEIRA, V.C.A.; SILVEIRA, N.F.A. , NASCIMENTO , M.D.D. and GOMES ,R.A.R
 المصدر:
MICROBIOLOGICAL EXAMINATION METHODS OF FOOD AND WATE A Laboratory Manual
المصدر:
MICROBIOLOGICAL EXAMINATION METHODS OF FOOD AND WATE A Laboratory Manual
 الجزء والصفحة:
الجزء والصفحة:
 17-3-2016
17-3-2016
 6597
6597




+
-
20
Pseudomonas
1. Introduction
The members of the genus Pseudomonas are bacteria exhibiting metabolic and nutritional characteristics extremely versatility, which allows them to utilize an enormous variety of organic compounds as source of carbon and energy. As a function of this versatility, they occupy and harbor in highly diversified ecological niches and are widely distributed in nature, in water and in foods.
2. Taxonomy
This genus has gone through great taxonomic changes, which began already in the 1st edition of Bergey’s Manual of Systematic Bacteriology (Krieg & Holt, 1984). In this edition, the species were divided into five groups, based on rRNA similarity. These five groups served further as a basis for later studies, which in turn led to the creation of various new genera, presented in Table 1.
Group I. Group I contained by far the largest number of species and, for that reason, the generic name of Pseudomonas was retained for this group.
Group II. The species of Group II (P. cepacia, P. mallei, P. pseudomallei, P. caryophylli, P. gladioli, P. pickettii e P. solonacearum) were transferred to the new genus Burkholderia. Later, P. pickettii and P. solonacearum were reallocated to the new genus Ralstonia.
Group III. The species belonging to Group III were subdivided into five genera: P. delafieldi and P. facilis were transferred to the new genus Acidovorax. P. flava, P. pseudoflava and P. palleronii were transferred to the new genus Hydrogenophaga. P saccharofila was transferred to the new genus Pelomonas. P. testosteroni and P. acidovorans were transferred to the new genus Comamonas, but later on P.acidovorans was reallocated to the new genus Delftia.
Group IV. The species of group IV (P. diminuta and P. vesiculares) were transferred to the new genus Brevundimonas.
Group V. The sole species of group V (P. maltophilia) was initially transferred to the genus Xanthomonas, but later on reallocated to the new genus Stenotrophomonas.
Other species. In addition to the species of the five groups, many other species were described in the 1st edi-tion of Bergey’s Manual of Systematic Bacteriology, but with uncertain affiliation and/or nomenclature. Many of these species have also been transferred to new genera or to already existing genera (Table 1).
Among the species reallocated into new genera, some are common in foods, including Shewanella putrefaciens, Janthinobacterium lividum and Stenotrophomonas maltophilia.
- Pseudomonas Migula 1894
Pseudomonas species are straight or slightly curved rods, Gram-negative, nonsporeforming and rarely nonmotile. Chemoorganotrophic, most species do not require organic growth factors and are able to grow in chemically minimally defined media, using one single compound as source of carbon, and ammonia or nitrate as source of nitrogen. Aerobic, having a strictly respiratory type of metabolism with oxygen as final electron acceptor; in some species nitrate can be used as an alternate electron acceptor, allowing growth to occur under anaerobic conditions. The optimal temperature of the majority of the species lies around 28ºC,with some species growing at 45ºC and several growing well at 4ºC (psychrotrophic).
Most species fail to grow under acid conditions (pH 4.5 or lower). Catalase-positive, oxidase-positive -or nega-tive (Palleroni, 2005a). Pigmentation is a characteristic common to many species. In fact, the colonies and other cell masses of Pseudomonas always display some colors due to normal cellular components, which, in some instances, become quite apparent. For example, P. stutzeri is a non-pigmented species but the colonies of many strains become dark brown due to high concentration of cytochrome c in the cells. P. aeruginosa and other fluorescent pseudomonads may produce blue, green or orange phenazines and green-yellow pyoverdines. The phenazine best known is pyocyanin (blue), which can be stimulated in P. aeruginosa using the King A Medium. P. chlororaphis subsp. chlororaphis produces chlororaphin (green) and P. chlororaphis subsp. aureofaciens produces phenazine-monocarboxylic acid (orange) (Palleroni, 2005a).
Table 1 Changes in the nomenclature of members of the genus Pseudomonas (Palleroni, 2005a, Euzéby, 2011).


Several species of Pseudomonas are pathogenic to plants and several are opportunistic pathogens to humans, being associated with infections in individuals with debilitated immune systems. According to Euzéby (2006) the most common in humans is Pseudomonas aeruginosa, but Pseudomonas alcaligenes, Pseudomonas balearica, Pseudomonas chlororaphis, Pseudomonas fluorescens, Pseudomonas mendocina, Pseudomonas monteilii, Pseudomonas mosselii, Pseudomonas putida, Pseudomonas stutzeri and Pseudomonas pseudoalcaligenes are also isolated from clinical specimens. After P. aeruginosa, the most frequent species are P. fluorescens, P. putida and P. stutzeri, although these species exhibit a lower degree of virulence and a more limited invasive power.
According to Euzéby (2006) Pseudomonas can be found in water (freshwater, brackish water and seawater), in soil, in suspended dust, in the air and in vegetables. Many strains are psychrotrophic and may alter or spoil food products, biological reagents, injectable solutes, blood and blood derivatives stored under refrigeration. Also as function of the richness of their metabolic pathways, they frequently are capable of resisting to the action of numerous antiseptics and antibiotics. This explains their increasingly more frequent presence in hospital environments, isolated from humid or moist places or items, such as sinks, siphons, washing & resting room clothes and objects, water containers, etc. They are rarely found on the skin or muscles of humans and animals, but are common as part of the intestinal flora.
Pseudomonas in treated water intended for human consumption: According to ICMSF (2000) Pseudomonas are common in raw water and are also found in treated water. The aim and purpose of treating raw water is the destruction of pathogens and the presence of some degree of viable microorganisms after the treatment is normal and acceptable. However, high counts at the points of consumption reflect growth or recontamination somewhere in the distribution system, that is, in the reservoirs and piping system. Growth may occur due to the survival and recovery of injured cells of native microorganisms, including Pseudomonas, Flavobacterium, Arthrobacter and Aeromonas. According to WHO (2011) P. aeruginosa can be significant in certain settings such as health-care facilities, but there is no evidence that normal uses of drinking-water supplies are a source of infection in the general population. However, the presence of high numbers of P. aeruginosa in potable water, notably in packaged water, can be associated with complaints about taste, odor and turbidity.
Pseudomonas in mineral water and natural water: At the point of emergency mineral water and natural water exhibit a normal microbiota between 10 and 100 CFU/ml, composed primarily by Pseudomonas, Flavobacterium, Moraxella, Acinetobacter and Xanthomonas. This natural microbiota is generally aerobic, has a very low nitrogen requirement and is able to grow at low temperatures, with small amounts of carbon com-pounds (ICMSF, 2000).
After bottling, multiplication of this microbiota is nor-mal and characterized by alternating increases and reductions in the counts. Immediately upon completion of filling, the population increases using the organic material on the surface of the packages and the oxygen dissolved during the filling operation. Twelve hours after filling, counts generally reach values ten-fold greater than normal at the point of emergence, but may reach values as high as 104–105 CFU/ml (on plates incubated at 20–22ºC/72 h).
The higher counts are more common in bottled water filled into plastic package, probably due to the fact that plastic is a surface that is more favorable to microbial growth (ICMSF, 2000). After this, the population declines due to the depletion of nutrients, but begins to grow again utilizing the organic material of the dead cells and so forth.
Several species of Pseudomonas have been isolated from mineral water, including P. mandelii, P. migulae, P. rhodesiae and P. veronii (Palleroni, 2005a), but the main concern is the presence of P. aeruginosa.
Pseudomonas in foods: The occurrence of Pseudomonas in foods is extremely common, and associated with the spoilage of meats and meat products, milk and dairy products, fish and seafood, eggs and vegetables. In raw poultry meat, kept at a temperature from minus 2ºC to 5ºC, the pseudomonads are the main spoilage microorganisms, because they have the lowest generation time at these low temperatures. Among the more frequently found species are P. fragi, P. fluorescens and P. putida (ICMSF, 2000).
In non-processed, cold-stored beef, the pseudomonads represent more than 50% of the deterioration-causing microbiota. P. fragi, P. lundensis and P. fluorescens are the most frequently encountered species. At the beginning of the spoilage process, they cause a fruity sweet odor, as a result of the formation of ethylic esters. In the more advanced stages, the production of sulphur compounds causes a putrid odor (ICMSF, 2000). In non-cured, cooked meat, packaged under normal atmosphere and refrigerated, Gram-negative bacteria, including Pseudomonas, are also the main spoilage agents (ICMSF, 2000).
In refrigerated pasteurized milk, the pseudomonads are part of the spoilage microbiota, although they do not survive pasteurization (post-processing recontamination). They do also not survive in sterilized milk, but their lipolytic and proteolytic enzymes may remain in the product and cause alterations. In heat-treated milk cream, post-processing recontamination by pseudomonads has a strong impact on the quality of the finished product (ICMSF, 2000). The most common species are P. fragi, P. fluorescens and P. taetrolens, in addition to P. synxantha in milk cream, where they produce a deep yellow-to-orange pigment (Palleroni, 1984, Palleroni, 2005a).
In whole raw eggs, the pseudomonads are the main cause of deterioration, because they generally are the first micro-organism to penetrate the egg shell, and are resistant to the natural antimicrobial agents of the egg white. Among the species found in eggs are P. putida (causes fluorescence of the egg white), P. fluorescens (causes a pink color to develop in the egg white), P. mucidolens and P. taetrolens (ICMSF, 2000, Palleroni, 1984, Palleroni, 2005a).
- Shewanella MacDonell & Colwell 1986
According to Bowman (2005) the species of the genus Shewanella are Gram-negative, rod-shaped, oxidase and catalase-positive bacteria. The colonies frequently have an orange-like, pinky, salmon or slightly tanned color, due to the accumulation of cytochrome. Facultative anaerobic; oxygen is used as final electron acceptor during aerobic growth and the anaerobic growth is predominantly respiratory (anaerobic respiration), using nitrate, nitrite, Fe3+, Mn3+, fumarate and various sulphur compounds as alternate electron acceptor. Trimethylamine N-oxide (TMAO) can also be used as final acceptor of electrons and its reduction to trimethylamine is generally responsible for the odor associated with the spoilage of foods caused by Shewanella. Three species also present fermentative metabolism, Shewanella frigidimarina and Shewanella benthica, which ferment glucose, and Shewanella gelidimarina, which ferments N-acetylglucosamine. The majority of the species grows at 4ºC and forms H2S from thiosulphate. Several species require NaCl for growth. Two species are among the most important food spoilage microorganisms, Shewanella putrefaciens and Shewanella baltica, found in dairy products, meat products, fish and seafood.
Shewanella putrefaciens (synonym Pseudomonas putrefaciens). According to Bowman (2005) S. putrefacies was described for the first time in 1931, as Achromobacter putrefaciens. In 1941, it was transferred to the genus Pseudomonas, because of rod-shaped morphology, motility and strictly respiratory, non-fermentative metabolism. Based on the mol% G+C value, lower than that of the pseudomonads, the species was transferred in 1971 to the genus Alteromonas. Finally, based on the rRNA 5S sequence, in 1986 it was transferred to the new genus Shewanella. The nomenclature of this species continues in evolution, since it is composed by a variety of heterogenic strains, which includes at least four DNA hybridization groups. The strains of Group IV were transferred to the new species Shewanella algae (in1990) and the strains of Group II to the new species Shewanella baltica (in 1998). The strains of Group I are considered Shewanella putrefaciens “sensu stricto” and the new taxonomic position of the strains belonging to Group III has not yet been defined so far. The main characteristics of these four groups are described in Table 2.
Table.2 Characteristics differentiating the strains of Shewanella putrefaciens “senso stricto”, Shewanella algae, Shewanella baltica and Shewanella putrefaciens Group III (Bowman, 2005).
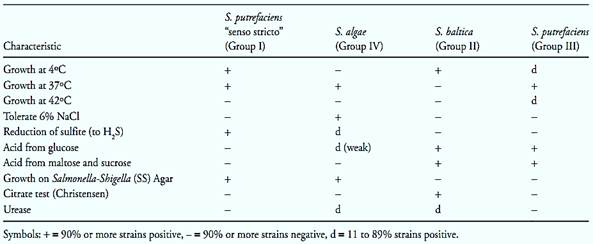
Colonies on Nutrient Agar are light tan to salmon pink, opaque, circular, convex with entire edges, and butyrous in consistency. NaCl is usually not required for growth and can tolerate up to 6% Does not require growth factors. Mesophilic, growth occurs between 10 and 40ºC, optimum, 30–35ºC. There are psychrotrophic strains that grow at 4ºC (Bowman, 2005).
According to ICMSF (2000) Shewanella putrefaciens is a common spoilage microorganism in raw poultry meat, in which it causes a sulphydric odor. It affects mainly meats with a pH greater than six, such as chicken leg cuts, the pH of which is higher than that of breast meat. In raw, vacuum-packed beef, Shewanella putrefaciens is a powerful producer of H2S and causes quick deterioration the products with higher pH values.
- Janthinobacterium De Ley et al. 1978 emend. Lincoln et al. 1999
According to Gillis and Logan (2005) the species of Janthinobacterium are Gram-negative rods, motile, oxidase and catalase-positive. Chemoorganotrophic, aerobic, has a strictly respiratory type of metabolism with oxygen as the final electron acceptor. Growth occurs on common peptone-based media. Citrate and ammonium ions can be used as sole carbon and nitrogen sources, respectively. Small amounts of acid, but no gas are produced from glucose and some other carbohydrates. Growth occurs from 4°C to about 30ºC, with an optimum at around 25ºC, and does not occur at 37ºC. The optimal pH is between 7 and 8 with no growth below pH 5. Many strains produce the violet pigment violacein, but strains producing non-pigmented or partially pigmented colonies are often encountered (strains producing partially pigmented colonies means a strain forming both pigmented and non-pigmented colonies on the same inoculated plate).
Janthinobacterium lividum (synonym Pseudomonas mephitica). Pseudomonas mephitica was the name initially proposed by Claydon & Hammer (1939) for a new bacterial species isolated from spoiled butter. The deterioration described by Claydon & Hammer (1939) was characterized by a stinking odor, such as that exhaled by the skunk (Mephitis mephitis). Anzai et al. (2000) reported a straight phylogenetic relationship between P. mephitica and Janthinobacterium lividum, a species discovered in 1891 (as Bacillus lividus) and also already called Chromobacterium lividum. The majority of the strains of J. lividum produce the violet violacein dye, but there are strains that, such as P. mephitica, are not pigmented. In 2008, based on the comparison of the rRNA 16S sequence and that of numerous phonotypical characteristics, P. mephitica was recognized as a synonym of J. lividum (Kämpfer et al., 2008).
The original description of Claydon & Hammer (1939) for P. mephitica: Gram negative rods, motile, nonsporeforming. Optimal temperature around 21ºC, grows slow at 5ºC and at 30ºC, does not grow at 37ºC. pH for growth between 5.0 and at 7.5. Acid not produced from lactose, slowly produced from maltose and glucose, with a tendency to revert. Not pigmented.
Gillis and Logan (2005) and Euzéby (2004) description for J. lividum, in addition to those which are common to all the species of the genus: Strict aerobe, does not grow under anaerobic conditions. More than 90% of the strains produce violacein (the strains previously denominated as P. mephitica do not produce the violacein pigment). The majority of the strains grow at 4ºC, but none at 37ºC. The majority does not grow in the presence of 2% NaCl or higher. The majority grows at pH 5.0, but only few at pH 4.0. The colonies have a gelatinous or rubber-like visual aspect and, in broth, the pigmented strains cause the formation of a violet ring on the surface. In agar, the violet pigmentation is generally little intense and appears only at a slow rate. About 50% of the strains grow in MacConkey Agar.
According to ICMSF (2000) Janthinobacterium lividum is isolated from the soil, freshwater, the marine environment and waste water. Contamination of food items by pigmented strains may cause the formation of blue stains. Just like the pseudomonas, this bacterium is one of the main spoilage microorganisms of cold-stored raw poultry meat. In raw beef, it may cause the pro-duction of H2S, and is sometimes accompanied by a greenish discoloration (Tompkin et al., 2001). In milk and dairy products it causes a stinking odor (Euzéby, 2004).
- Stenotrophomonas Palleroni & Bradbury 1993
This genus was created by Palleroni and Bradbury (1993) to accommodate Pseudomonas maltophilia and during several years S. maltophilia was the sole species of the genus, which later received two new species.
According to Palleroni (2005b) the stenotrophomonas are Gram-negative rods, nonsporeforming, motile, cata-lase and gelatinase-positive. Cytochrome c is absent and oxidase is negative (in the original description of the new genus Stenotrophomonas by Palleroni & Bradbury, 1993, the oxidase reaction was wrongly described as positive). Chemoorganotrophic, aerobic, has a strictly respiratory type of metabolism with oxygen as the final electron acceptor. Nitrate is reduced to nitrite but is not used as sole source of nitrogen for growth. The colonies are yellow, greenish or gray, which may turn brownish with incubation time. No growth occurs at 4ºC or 41ºC; the optimum growth temperature is around 35ºC. They exhibit strong lipolytic activity, characterized by the hydrolysis of Tween 80. Growth factors required, mainly methionine.
Stenotrophomonas maltophilia (synonym Pseudomonas maltophilia). According to Palleroni (2005b) S. maltophilia is nutritionally not as versatile as the pseudomonads, a fact that has served as the basis for denominating the new genus Stenotrophomonas (from Greek, means a unit that is able to utilize few substrates). This species is typically capable of using disaccharides (maltose, lactose, cellobiose) as sole sources of carbon and energy, a feature that is also rare among the pseudomonads. Growth in lactose is poorer, prob-ably because it does not utilize the galactose unit. Acid is produced from maltose, but not from glucose. The colonies may be yellow or greenish yellow in several culture media, producing pigments that do not diffuse into the medium, probably flavins. Some strains develop a brown discoloration in agar media.
Opportunistic pathogen, S. maltophilia has been associated to various types of infection and has been isolated from contaminated tissue cultures, streptomycin solutions, distilled water, incubator reservoirs, respirators, nebulizers, and evacuated blood collection tubes. Common in the rhizosphere of several cultivated plants, found in water and in foods such as raw and pasteurized milk, frozen fish and rotten eggs (Palleroni, 2005b).
References
Silva, N.D .; Taniwaki, M.H. ; Junqueira, V.C.A.; Silveira, N.F.A. , Nasdcimento , M.D.D. and Gomes ,R.A.R .(2013) . Microbiological examination methods of food and water a laboratory Manual. Institute of Food Technology – ITAL, Campinas, SP, Brazil .
Anzai, Y., Kim, H., Park, J.Y., Wakabayashi, H. & Oyaizu, H. (2000) Phylogenetic affiliation of the pseudomonads based on 16S rRNA sequence. International Journal of Systematic and Evolutionary Microbiology, 50, 1563–1589.
Bowman, J.P. (2005) Genus Shewanella. In: Brenner, D.J., Krieg, N.R. & Staley, J.T. (eds). Bergey’s Manual of Systematic Bacteriology. Volume 2, Part B. 2nd edition. New York, Springer Science+Business Media Inc. pp. 480–491.
Claydon, T.J. & Hammer, B.W., 1939. A skunk-like odor of bacterial origin in butter. Journal of Bacteriology, 37, 251–258. Euzéby, J.P. (2004) Janthinobacterium, Janthinobacterium lividum. Euzéby, J.P. Dictionnaire de Bactériologie Vétérinaire. [Online] France. Available from: http://www.bacterio.cict.fr/bacdico/jj/janthinobacterium.html [Accessed 24th October 2011].
Euzéby, J.P. (2006) Pseudomonadales, Pseudomonadaceae, Pseudomonas. In: Euzéby, J.P. Dictionnaire de Bactériologie Vétérinaire. [Online] France. Available from: http://www.bac-terio.cict.fr/bacdico/pp/pseudomonadales.html [Accessed 24th October 2011].
Euzéby, J.P. (2011) List of Prokaryotic Names with Standing in Nomenclature. [Online] Available from: http://www.bacterio. cict.fr/ [Accessed 24th October 2011].
Gillis, M. & Logan, N.A. (2005) Genus Janthinobacterium. In: Brenner, D.J., Krieg, N.R. & Staley, J.T. (eds). Bergey’s Manual of Systematic Bacteriology. Volume 2, Part C. 2nd edition. New York, Springer Science+Business Media Inc. pp. 636–642.
Hunt, M.E. & Rice, E.W. (2005) Microbiological examination. In: Eaton, A.D., Clesceri, L.S., Rice, E.W. & Greenberg, A.E. (eds). Standard Methods for the Examination of Water & Wastewater. 21st edition. Washington, American Public Health Association (APHA), American Water Works Association (AWWA) & Water Environment Federation (WEF). Part 9213, pp. 9.33–9.34.
ICMSF (International Commission on Microbiological Specifications for Foods) (2000) Microorganisms in Foods 6 – Microbial Ecology of Food Commodities. Gaithersburg, Maryland, Aspen Publishers.
International Organization for Standardization (2010) ISO 6887–5:2010. Microbiology of food and animal feeding stuffs - Preparation of test samples, initial suspension and decimal dilutions for microbiological examination - Part 5: Specific rules for the preparation of milk and milk products. Geneva, ISO.
International Organization for Standardization (2007) ISO 7218:2007. Microbiology of food and animal stuffs – General requirements and guidance for microbiological examinations. Geneva, ISO.
International Organization for Standardization (2009) ISO 11059:2009. Milk and milk products – Method for the enumeration of Pseudomonas spp. Geneva, ISO.
International Organization for Standardization (2010) ISO 13720:2010. Meat and meat products – Enumeration of presumptive Pseudomonas spp. Geneva, ISO.
International Organization for Standardization (2006) ISO 16266:2006. Water quality – Detection and enumeration of Pseudomonas aeruginosa – Method by membrane filtration. Geneva, ISO.
Kämpfer, P., Falsen, E. & Busse, H.J. (2008) Reclassification of Pseudomonas mephitica Claydon and Hammer 1939 as a later heterotypic synonym of Janthinobacterium lividum (Eisenberg 1891) De Ley et al. 1978. International Journal of Systematic and Evolutionary Microbiology, 58, 136–138.
Palleroni, N.J. (1984) Genus I. Pseudomonas. In: Krieg, N.R. & Holt, J.G. (eds). Bergey’s Manual of Systematic Bacteriology. 1st edition. Volume 1. Baltimore: Williams & Wilkins.
Palleroni, N.J. (2005a) Genus Pseudomonas. In: Brenner, D.J., Krieg, N.R. & Staley, J.T. (eds). Bergey’s Manual of Systematic Bacteriology. Volume 2, Part B. 2nd edition. New York, Springer Science+Business Media Inc. pp. 323–379.
Palleroni, N.J. (2005b) Genus Stenotrophomonas. In: Brenner, D.J., Krieg, N.R. & Staley, J.T. (eds). Bergey’s Manual of Systematic Bacteriology. Volume 2, Part B. 2nd edition. New York, Springer Science+Business Media Inc. pp. 107–115.
Palleroni, N.J. & Bradbury, J.F. (1993). Stenotrophomonas, a new bacterial genus for Xanthomonas maltophilia (Hugh 1980) Swings et al. 1983. International Journal of Systematic Bacteriology, 43, 606–609.
Tompkin, R.B., McNamara, A.M. & Acuff, G.R. (2001) Meat and meat products. In: Downes, F.P. & Ito, K. (eds). Compendium of Methods for the Microbiological Examination of Foods. 4th edition. Washington, American Public Health Association. Chapter 45, pp. 463–471.
WHO (ed.) (2011) Guidelines for drinking-water quality. 4th edi-tion. [Online] Geneva, Switzerland, World Health Organization. Chapter 11 – Microbial Fact sheets, pp. 249–250. Available from: http://www.who.int/water_sanitation_health/publications/2011/9789241548151_ch11.pdf [Accessed 14th December 2011].
 الاكثر قراءة في البكتيريا
الاكثر قراءة في البكتيريا
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















