























الفاكهة والاشجار المثمرة
نخيل التمر

النخيل والتمور
آفات وامراض النخيل وطرق مكافحتها
التفاح
الرمان
التين
اشجار القشطة
الافو كادو او الزبدية
البشمله او الاكي دنيا
التوت
التين الشوكي
الجوز
الزيتون
السفرجل
العنب او الكرمة
الفستق
الكاكي او الخرما او الخرمالو
الكمثري(الاجاص)
المانجو
الموز
النبق او السدر
فاكة البابايا او الباباظ
الكيوي
الحمضيات
آفات وامراض الحمضيات
مقالات منوعة عن الحمضيات
الاشجار ذات النواة الحجرية
الاجاص او البرقوق
الخوخ
الكرز
المشمش
الدراق
مواضيع عامة
اللوز
الفراولة او الشليك
الجوافة
الخروب(الخرنوب)
الاناناس
مواضيع متنوعة عن اشجار الفاكهة
التمر هندي
الكستناء
شجرة البيكان ( البيقان )
البندق
المحاصيل
المحاصيل البقولية
الباقلاء (الفول)
الحمص
الترمس
العدس
الماش
اللوبياء
الفاصولياء
مواضيع متنوعة عن البقوليات
فاصوليا الليما والسيفا
محاصيل الاعلاف و المراعي
محاصيل الالياف
القطن
الكتان
القنب
الجوت و الجلجل
محصول الرامي
محصول السيسال
مواضيع متنوعة عن محاصيل الألياف
محاصيل زيتية
السمسم
فستق الحقل
فول الصويا
عباد الشمس (دوار الشمس)
العصفر (القرطم)
السلجم ( اللفت الزيتي )
مواضيع متنوعة عن المحاصيل الزيتية
الخروع
محاصيل الحبوب
الذرة
محصول الرز
محصول القمح
محصول الشعير
الشيلم
الشوفان (الهرطمان)
الدخن
محاصيل الخضر
الباذنجان
الطماطم
البطاطس(البطاطا)
محصول الفلفل
محصول الخس
البصل
الثوم
القرعيات
الخيار
الرقي (البطيخ الاحمر)
البطيخ
آفات وامراض القرعيات
مواضيع متنوعة عن القرعيات
البازلاء اوالبسلة
مواضيع متنوعة عن الخضر
الملفوف ( اللهانة او الكرنب )
القرنبيط او القرنابيط
اللفت ( الشلغم )
الفجل
السبانخ
الخرشوف ( الارضي شوكي )
الكرفس
القلقاس
الجزر
البطاطا الحلوه
القرع
الباميه
البروكلي او القرنابيط الأخضر
البنجر او الشمندر او الشوندر
عيش الغراب او المشروم او الأفطر
المحاصيل المنبهة و المحاصيل المخدرة
مواضيع متنوعة عن المحاصيل المنبهة
التبغ
التنباك
الشاي
البن ( القهوة )
المحاصيل السكرية
قصب السكر
بنجر السكر
مواضيع متنوعة عن المحاصيل
نباتات الزينة والنباتات الطبية والعطرية
نباتات الزينة
النباتات الطبية والعطرية
الحشرات النافعة
النحل
نحل العسل
عسل النحل ومنتجات النحل الاخرى
آفات وامراض النحل
دودة القز(الحرير)
آفات وامراض دودة الحرير
تربية ديدان الحرير وانتاج الحرير الطبيعي
تقنيات زراعية
الاسمدة
الزراعة العضوية
الزراعة النسيجية
الزراعة بدون تربة
الزراعة المحمية
المبيدات الزراعية
انظمة الري الحديثة
التصنيع الزراعي
تصنيع الاعلاف
صناعات غذائية
حفظ الاغذية
الانتاج الحيواني
الطيور الداجنة
الدواجن
دجاج البيض
دجاج اللحم
امراض الدواجن
الاسماك
الاسماك
الامراض التي تصيب الاسماك
الابقار والجاموس
الابقار
الجاموس
امراض الابقار والجاموس
الاغنام
الاغنام والماعز
الامراض التي تصيب الاغنام والماعز
آفات وامراض النبات وطرق مكافحتها
الحشرات
الحشرات الطبية و البيطرية
طرق ووسائل مكافحة الحشرات
الصفات الخارجية والتركيب التشريحي للحشرات
مواضيع متنوعة عن الحشرات
انواع واجناس الحشرات الضارة بالنبات
المراتب التصنيفية للحشرات
امراض النبات ومسبباتها
الفطريات والامراض التي تسببها للنبات
البكتريا والامراض التي تسببها للنبات
الفايروسات والامراض التي تسببها للنبات
الاكاروسات (الحلم)
الديدان الثعبانية (النيماتودا)
امراض النبات غير الطفيلية (الفسيولوجية) وامراض النبات الناتجة عن بعض العناصر
مواضيع متنوعة عن امراض النبات ومسبباتها
الحشائش والنباتات الضارة
الحشائش والنباتات المتطفلة
طرق ووسائل مكافحة الحشائش والنباتات المتطفلة
آفات المواد المخزونة
مواضيع متنوعة عن آفات النبات
مواضيع متنوعة عن الزراعة
المكائن والالات الزراعية

التغيرات الفسيولوجية السابقة للإزهار في الحمضيات
 المؤلف:
أ.د مصطفى عاطف الحمادي واخرون
المؤلف:
أ.د مصطفى عاطف الحمادي واخرون
 المصدر:
الموالح (الإنتاج والتحسين الوراثي)
المصدر:
الموالح (الإنتاج والتحسين الوراثي)
 الجزء والصفحة:
ص 425-443
الجزء والصفحة:
ص 425-443
 14-8-2022
14-8-2022
 3803
3803




+
-
20
التغيرات الفسيولوجية السابقة للإزهار في الحمضيات
أجريت العديد من الدراسات لتحديد العوامل الفسيولوجية التي تتحكم في إزهار الموالح (1990 ,Davenport) والعوامل الأكثر احتمالا لهذا التحكم هي الكربوهيدرات والهرمونات، ودرجة الحرارة، والعلاقات المائية والغذائية. والنظرية التي تقترح تحكم الكربوهيدرات في الإزهار تبني على أساس حقيقة أن الفرع أو الجذع عند تحليقه يزيد تحفيز الإزهار وعقد الثمار ومحتوى النشا في الأفرع، وقد يحدث ذلك نظرا لأن التحليق يثبط نقل الكربوهيدرات في اللحاء إلى الجذور. وعلى العكس من ذلك فهناك العديد من الدراسات التي لم تجد علاقة ارتباط بين مستوى النشا في الأوراق أو الأغصان وحدوث الأزهار في الموالح (1990 ,Osland and Davenport, 1987, Davenport) مع أنه قد وجد أن مستويات الكربوهيدرات في الجذر تكون في بعض الأحوال مرتبطة مع الإزهار في تبادل الحمل في اليوسفي حيث ارتبط مستوى الكربوهيدرات المنخفض جدا في الجذور (نظرا للحمل الزائد) مع الإنتاج المحدود للأفرع والإزهار، وقد لوحظ ذلك بصفة خاصة في اليوسفي صنف Murcott والذي يحمل محصولا غزيرا في سنه الحمل الغزير لدرجة أن تنخفض كمية الكربوهيدرات في الجذور وتصير آثارا تقريبا وقد تموت الأشجار نتيجة لذلك (Murcott collapse)، وعلى ذلك يبدو أنه لابد من توافر مستوى حرج من الكربوهيدرات في الجذور.
وقد درس 1990 ,Davenport تحكم الهرمونات في الإزهار حيث تم معاملة الأفرع ببعض المواد الهرمونية الطبيعية وتقييم تأثير ذلك على الإزهار ووجد أن الرش بحامض الجبرليك (GA3) لأفرع الموالح قبل حدوث التحفيز الزهري يثبط الإزهار، لذلك فإن حامض الجبرليك يبدو أنه منظم لبعض الخطوات في عملية الأزهار (1964 ,Monselive and Halevy). ولكن دلت الدراسات علي التغيرات في المحتوى الداخلي للجبرلين بعدم وجود علاقة مؤكدة ما بين GA3 ونوعية البراعم (زهري أو خضري)، وقد يكون للجبرلين تأثير في الأزهار عن طريق عملية وسطية (1990 ,Davenport). ولكن لتأكيد ذلك يلزم مزيد من الدراسات البحثية.
وترتبط الحالة الغذائية للأشجار بطريقة مباشرة وغير مباشرة مع الأزهار في أشجار الموالح حيث وجد أن المستويات المرتفعة من النتروجين (وخاصة في أشجار الموالح الصغيرة السن) تحفز النمو الخضري ولا تنتج أزهارا، وعلى العكس من ذلك فإن المستوى المنخفض من النتروجين في الأوراق يشجع الإزهار ولكن تنخفض نسبة عقد الثمار وبالتالي كمية المحصول تحت هذه الظروف، حيث وجد أن الأشجار التي تعاني من النقص الشديد في النتروجين تنتج عددا قليلا من الأزهار، لذلك فالمحافظة على مستوى النتروجين في الورقة في الحدود المناسبة (2.5 - 2.7 %) تؤدي إلي إنتاج عددا متوسطا من الأزهار ويتم العقد فيها بنسبة كبيرة وبالتالي محصولا جيدا. وقد يؤثر النتروجين على صورة أمونيا على الإزهار مباشرة عن طريق تنظيم مستوى الأمونيا والبولي أمين Polyamine في البراعم (1988 ,Lovatt et al). وقد وجد أن الإجهاد المائي والحرارة المنخفضة يزيدان من محتوى الأوراق من الأمونيا وبالتالي الأزهار، كما وجد في كاليفورنيا أن الرش باليوريا (أو إضافتها) في فترة انخفاض درجة الحرارة في الشتاء للبرتقال أبو سره واشنطن أدت إلى زيادة محتوى الأوراق والبراعم من الأمونيا وعدد الأزهار / الشجرة وكان عدد الأزهار الناتجة مرتبطة ارتباطا موجبا مع طول فترة التحفيز بالحرارة المنخفضة. كما أوضح (1992 ,Rabe and Vender Walt) أن رش أشجار البرتقال الشموتي باليوريا بتركيز 1% قبل التزهير بفترة من 6 إلى 8 أسابيع أدت إلى زيادة الأزهار والمحصول.
ينشط تكوين البراعم الزهرية عند تعرض الأشجار لفترات من البرودة أو الإجهاد المائي، وتتميز البراعم الزهرية وتنمو عند دفء الجو أو توفر الماء الأرضي وإنهاء الإجهاد المائي، كما تؤثر درجات الحرارة على موعد الإزهار، كما أن للهرمونات النباتية دورا هاما في التحكم في فسيولوجيا إزهار الموالح، فقد وجد أن الجبرلين (GA3) فقط هو الذي له دور مثبط على تحول البراعم الزهرية، وقد يكون من أسباب صعوبة فهم العوامل التي تتحكم في إزهار الموالح جزئيا على سبيل المثال أنه يمكن تحفيز بعض البراعم بسهولة للتزهير بينما البعض الآخر (وحتى على نفس الفرع) يتطلب التعرض لظروف إجهاد أكثر شدة لتحفيزها، ومن الواضح أن العوامل المؤثرة على فسيولوجيا الإزهار في الموالح تحتاج لمزيد من الدراسات البحثية، ويتم الإزهار في الموالح على عدة مراحل كما يلي:
أ- مرحلة التحفيز الزهري (الدفع الزهري) Flower buds Induction
يقصد بالتحفيز الزهري حدوث تغيرات بيوكيميائية في البراعم وبالتالي تتحول البراعم الخضرية إلى براعم زهرية وهذا الطور غير مرئي ويشمل التغيرات التي تؤدي إلى التحول من النمو الخضري إلى الإنتاج الزهري (1990 ,Davenport)، ويبدأ التحفيز لإنتاج الإزهار بتوقف النمو الخضري أثناء السكون في الشتاء في البيئات التحت استوائية أو الأوقات الجافة في المناطق الاستوائية، ويبدو أن تعرض البراعم لدرجات حرارة أقل من 25 م لعدة أسابيع يكون مطلوبا لتحفيز تحول البراعم الخضرية إلى البراعم الزهرية بكميات مؤكدة (1990 ,Inoue) وفترات العطش لمدة أطول من 30 يوما تحت ظروف الحقل تدفع عدد كبير من البراعم للإزهار. وترتبط درجة التحفيز بمقدار الإجهاد وحدته واستمراره (1986, Southwick & Davenport). وعادة يتوقف النمو الخضري للأشجار الناضجة ويقل معدل نمو الجذور بانخفاض درجة الحرارة في الشتاء حتى ولو كانت الحرارة أعلى من 12.5 م، وأثناء هذه الفترة يبدأ حدوث بعض التغيرات البيوكيميائية في البراعم الخضرية وذلك للبدء في مرحلة التحفيز الزهري Flower buds Induction، وقد اقترح ( 1992 , Garcia- Luis et al ) أن نشوء البرعم Bud initiation قد يسبق عملية التحفيز، وعملية التحفيز الزهري في البراعم مرحلة غير عكسية أي أن البرعم لا يرتد ثانية إلى الحالة الخضرية (1987 ,Lord and Echard)، ويختلف موعد تحفيز البراعم والإزهار كثيرا من موسم إلى آخر وذلك طبقا لمعدلات درجة الحرارة وتوفر المياه ويجب الأخذ في الاعتبار أنه بالرغم من أن التعرض للبرودة والعطش يحفز التحول الزهري ولكنة ليس ضروريا بمعنى أن الأشجار يمكنها الإزهار حتى ولم تتعرض لأي منهما.
وقد استخدم الإجهاد المائي كطريقة عملية الدفع الأشجار للإزهار في مواعيد الإزهار التي يرغبها المنتج والتي تحتاجها الأسواق في بعض أنواع الموالح مثل الليمون الأضاليا والليمون البلدي المالح والترنح والتي تتميز بإزهارها
طوال العام طالما كانت الظروف البيئية مناسبة كما يلي:
• في إيطاليا يمنع الماء عن أشجار الليمون الأضاليا أثناء الصيف حتى تصبح الأشجار مجهده بشدة وفي أثناء هذه الفترة تحفز البراعم الزهرية ولكنها نادرا ما تنمو. ثم تروى الأشجار لدفعها للأزهار في الخريف. وبذلك ينتج محصول في الصيف التالي (Verelli" lemons")، وبصفة عامة تزهر الأشجار بعد 3-4 أسابيع بعد الري.
• في مصر يتم تصويم الليمون المالح للحصول على محصول غير موسمي، والتصويم عبارة عن تعطيش الأشجار لفترة من السنة ثم ريها فتزهر بعد الري أو بمعني آخر هو تنظيم ري الأشجار لدفعها للإزهار في الميعاد المرغوب، ويتم التصويم بطريقتين على النحو التالي:
1- الصيام الصغير:
وتتبع مع الأشجار التي لا يقل عمرها عن ست سنوات. وذلك بمنع الري عنها لمدة تتراوح من شهر ونصف إلى شهرين في السنة (يوليه وأغسطس) على حسب نوع التربة والظروف الجوية. ثم تروى الأشجار في شهر سبتمبر (بعد فترة التصويم) بدون إضافة أي أسمدة كيماوية، ثم تضاف معدلات الأسمدة النتروجينية والبوتاسية التي كانت ستضاف للأشجار في شهر مارس قبل الرية الثانية من التصويم ويؤدي ذلك إلى إزهار الأشجار وعقد الثمار ونموها قليلا ثم يتوقف نمو الثمار خلال أشهر الشتاء بسبب البرودة، ومع ارتفاع درجات الحرارة في الربيع تستأنف الثمار نموها وتصبح صالحة للقطف خلال الفترة من مارس وحتى يونيه، ويباع المحصول غير الموسمي بأسعار مرتفعة نظرا لعدم توفر إنتاج من ثمار الليمون المالح خلال هذه الفترة، وبذلك تزهر وتثمر الأشجار في حالة التصويم الصغير مرتان في العام.
2- الصيام الكبير:
المعاملة الصيام الكبير مع الأشجار الكبيرة التي لا يقل عمرها عن عشر سنوات ومنزرعة في أرض طميية عميقة، وفي هذه الطريقة يتم منع الري عن الأشجار لمدة 9 شهور في السنة وتروى لمدة ثلاث شهور فقط هي سبتمبر وأكتوبر ونوفمبر وبذلك تروي الأشجار ثلاث أو أربع ريات في السنة، وتزهر الأشجار في شهر سبتمبر وتعقد ثمارها ثم تنمو قليلا في شهري أكتوبر ونوفمبر ثم يبطؤ نموها أثناء الشتاء بسبب البرودة ويستمر توقف نمو الثمار خلال فصلي الربيع والصيف بسبب العطش نتيجة لعدم الري وتسمى الثمار في هذه الحالة بالثمار اللاصقة، وعندما تروي الأشجار ثانية في أشهر سبتمبر وأكتوبر ونوفمبر من العام التالي تعاود الثمار نموها وتكبر في الحجم وفي نفس الوقت تزهر الأشجار نتيجة لمعاودة الري يعد فترة طويلة من العطش، وتكون الثمار صالحة للقطف خلال الفترة من نوفمبر إلي فبراير، ويلاحظ أن الأشجار المعاملة بهذه الطريقة قد تزهر إزهارا عاديا في الربيع ولكن معظم هذه الأزهار تتساقط نتيجة للعطش الشديد الذي تعاني منه الأشجار المصومة، ونظرا لأن معاملة الصيام الكبير تعتبر معاملة قاسية فإنها لا تتبع إلا مع الأشجار القوية الكبيرة التي لها مجموع جذري كبير قريب من مستوي الماء الأرضي، كما أن هذه المعاملة قد تؤدي إلى موت بعض الأشجار نتيجة منع الري لفترة طويلة مما أدي بالمزارعين إلى التوقف عن إتباع هذه الطريقة الآن ويستخدمون طريقة التصويم الصغير فقط للحصول على محصول غير موسمي من الليمون المالح.
وباستخدام التصويم أمكن إمداد الأسواق بالليمون طوال العام كما هو مبين بالجدول التالي:
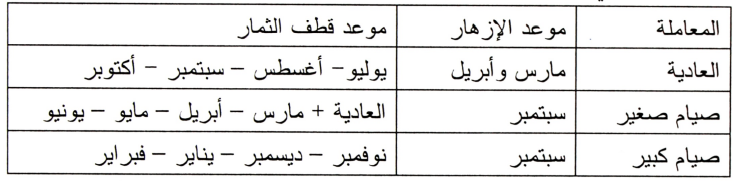
ومن حيث جودة الثمار فالثمار الناتجة من أشجار غير مصومة أفضل من ثمار الأشجار المصومة، كما أن الثمار الناتجة عن الصيام الصغير أفضل من تلك الناتجة من الصيام الكبير.
- وقد استخدمت هذه الطريقة أيضا لدفع الإزهار خارج الموسم في إسرائيل واسبانيا لليمون الأضاليا وفي فلوريدا بالولايات المتحدة لليمون التاهيتي.
- وقد وجد أن المعاملة بالجبرلين أثناء مرحلة التحفيز الزهري Flower buds Induction تؤدي إلى منع هذه العملية وبذلك تثبط عملية تحول البراعم الخضرية إلى براعم زهرية وبالتالي منع الإزهار خلال فترة ما (1990 ,Monselive and Halvey, 1964, Davenport) ثم عن طريق بعض المعاملات الأخرى مثل الإجهاد المائي أو المعاملة ببعض منظمات النمو المعيقة أو المؤخرة للنمو كطرق لدفع الأشجار للإزهار في مواعيد الإزهار التي يرغبها المنتج.
ب- مرحلة التميز والتكشف الزهري
Differentiation and Flower buds Initiation
تشمل هذه المرحلة التغيرات الهستولوجية والمورفولوجية في المرستيم الخضري ليصبح مرستيما زهريا (1990 ,Davenport) حيث يتفلطح المرستيم ويصبح منبسطا بدلا من الشكل القمي المدبب شكل (1، 2). ويبدأ تميز الأجزاء بتكون مبادئ السبلات يتبعه تكون الكرابل شكل (3).
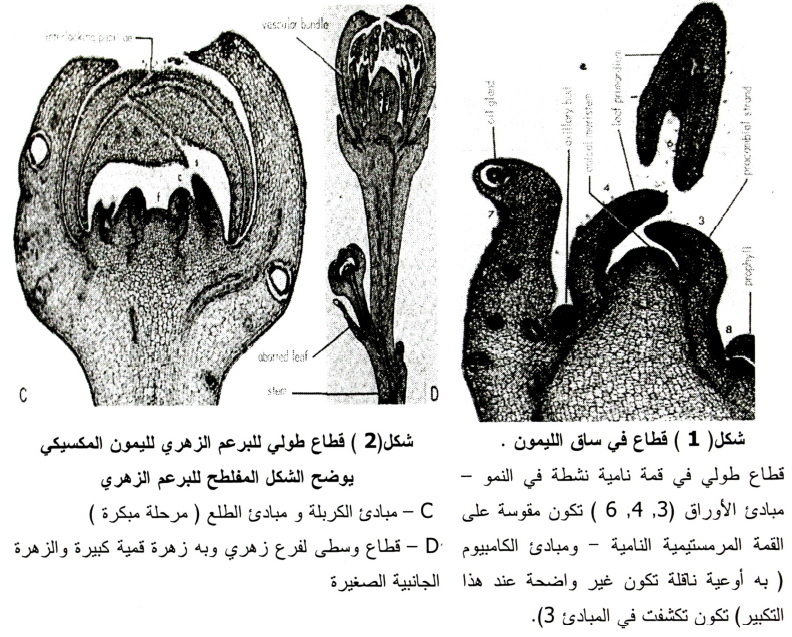
وتحدد الحالة التشريحية للقمة تتابع Sequence ومصير Disposition البراعم الجانبية (1987 ,Lord and Echard). فإذا كونت القمة النامية سبلات فإن البراعم الجانبية ستكون أزهار أيضا وعلى العكس إذا كونت القمة أوراق فإن البراعم الجانبية ستكون أزهار أيضا، وعلى العكس إذا كونت القمة أوراق فإن البراعم الجانبية ستكون أشواك. ولا يعتمد معدل تكوين الزهرة من توقيت خروج البرعم إلى الأزهار على موقع الزهرة أو نوع النورة ولكن يكون مرتبطا ارتباطا موجبا بعدد الوحدات الحرارية (1984 ,Lovatt et al). ويمكن توصيف المراحل المختلفة لنشأة وتطور الأجزاء الزهرية في الموالح فيما يلي:
1. نشأة وتطور المتاع:
أ- الكربلة والبلاسنتا:
يتكون المتاع في الموالح من حوالي عشرة وحدات (كرابل) كل منها يشبه قرن البسلة في مقطعه التركيبي والتي تتصل ببعضها بمحور رئيسي وسطي التكوين متاع مركب، ويتكون المتاع في الزهرة الناضجة من المبيض والميسم وقلم مخروطي عادة بينهما (شكل 3 - A&B)، وفي المراحل المبكرة جدا (شكل 4) فإن المتاع لا يكون مقفل من القمة ولكن يتكون من جدار دائري أو حلقة وبروزات داخل هذه الحلقة وهذه الحلقة تتكون من الكرابل المتلاصقة والتي تتكون في طورها المبدئي Primordial كتراكيب متجمعة Whori بأشكال هلالية Crescent shape. وتلاحظ الأشعة النخاعية في القطاع الطولي للزهرة بعد صبغه بالسفرانيين (شكل 3-B). وتنمو الكرابل إلى أعلا وتتجه حوافها إلى الداخل لتقابل النتوءات الداخلية الوسطية التي سبق ذكرها (شكل 4) وفي نفس الوقت فإن هذه الامتدادات الوسطية تنمو إلى أعلا لتكون محور الثمرة Core وتتحد مع حواف الكرابل، وعلى هذا فكل كربلة تحيط بفص Locule من المبيض (شكل 3- د). وتتكون البلاسنتا على الزاوية الداخلية لكل فص (وهذه هي منطقة أنسجة مستغلظة والتي تحمل البويضات)، وقد أوضح (1942 ,Ford) أن البويضات تنشأ من حواف الكرابل.
وتوجد قناة في القلم مغطاة من الداخل بخلايا من القشرة (1942 , Banerji & Ford 195) تمتد من البلاسنتا لكل كربلة إلى فتحة ضيقة على سطح الميسم. وتنمو الأنابيب اللقاحية من خلال هذه القنوات متجهة إلى البويضات، ولكن ذكر (1954 ,Banerji) أن الأنبوبة اللقاحية تنمو بين الخلايا ولا تنمو خلال هذه القناة.
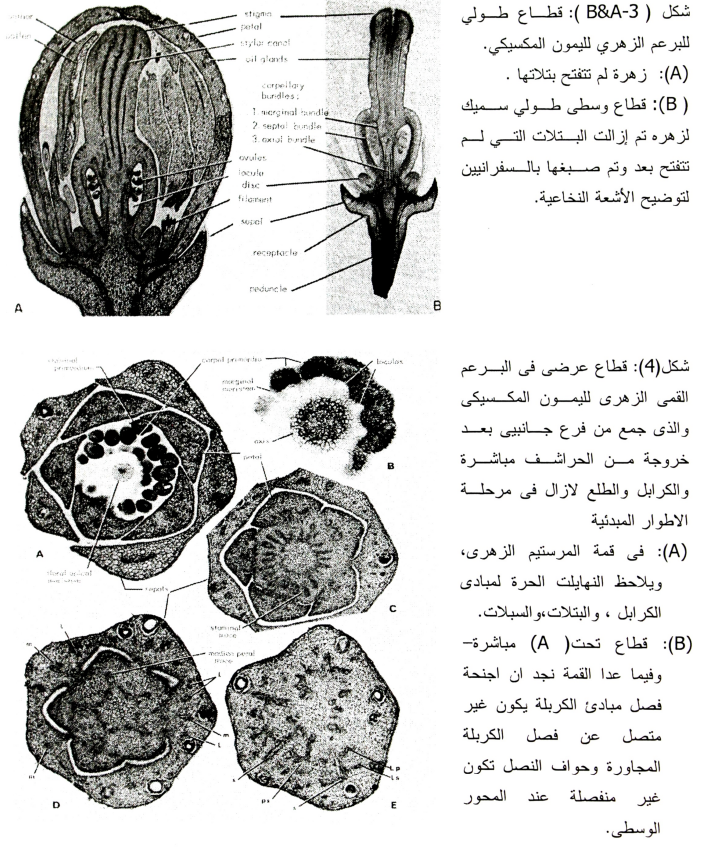
ب- البويضات:
تنشا البويضات (شكل 5) عندما تكون الازهار لازالت صغيرة جدا كنموات خارجية من البلاسنتا وهذا النمو الخارجي يشمل العديد من الصفوف الخلوية، وتبدأ البويضة في النمو إلى أعلى بسرعة لتأخذ في النهاية شكل بيضي مقلوب Anatropous وتتكون البويضة الناضجة من حامل Funiculus وكتلة وسطية من الخلايا (النيوسيلة) والكيس الجنيني في داخل النيوسيلة والغلافين Integuments المحيطان بالنيوسيلة، ويتواجد في قمة النيوسيلة فتحة خلال الغطاءين السابقين وهذه الفتحة عبارة عن الميكروبيل، وفي البويضة المقلوبة Anatropous تكون الناحية البعيدة عن البلاسنتا عبارة عن منطقة الكلازا.
ج- تكوين الكيس الجنيني والمتاع عامة:
تتميز خلية من الطبقة الثانية للخلايا قرب النيوسيلة (وذلك قبل تكون الأغلفة Integument) ويمكن تمييز هذه الخلية بحجمها الكبير والنواة الكبيرة (شكل 5- A). وهذه هي الخلية الأمية الجرثومية Archasporial cell، وتنقسم هذه الخلية مره (مكونة خليتين يتكون بينهما جدار أفقي) وبذلك تبدأ الخلية الخارجية منها بتكوين النسيج المغذي Tapetal Cell وتكون الخلية الداخلية الخلية الأمية للكيس الجنيني Megasporocyte (شكل 5- B).
وتنقسم الخلية العليا (Tapetal) لتكون ثمانية صفوف من الخلايا وتصبح الخلية الأمية للكيس الجنيني مدفونة في نسيج في وسط النيوسيلة. ويوضح شكل (5 C&D) تكوين خلايا النسيج المغذى وكذا الأغلفة Integuments متكونة جزئيا وقد لوحظ أنه في بعض الأحيان يتكون أكثر من خلية أمية جرثومية واحدة في المبيض ولذا يتواجد أكثر من كيس جنيني واحد في المبيض (1954 ,Banerji).
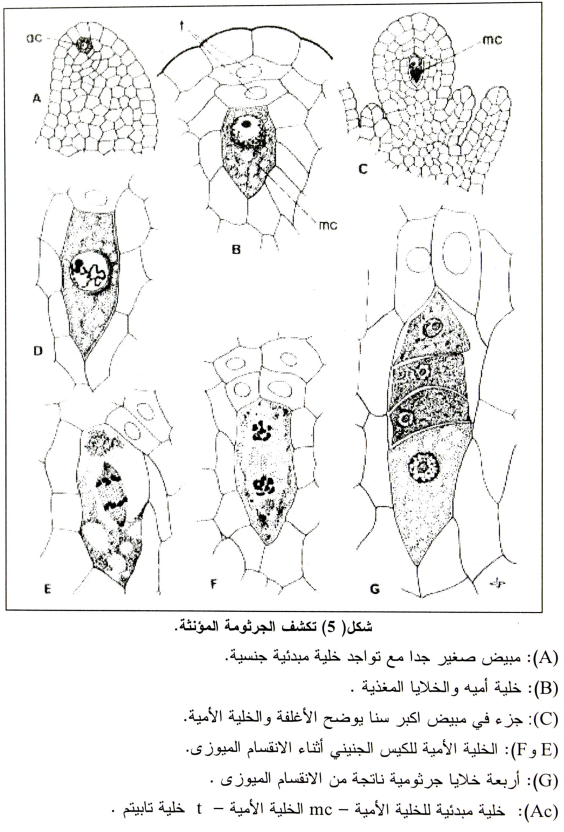
د- الانقسام الاختزالي وتكون الجاميطات المؤنثة:
تنمو الخلية الجرثومية الأمية للكيس الجنيني إلى أضعاف حجمها وتصبح مستطيلة (شكل 5). وتبدأ عملية تزاوج الكروموزومات المتجانسة في مرحلة Prophase للانقسام الأول، ويحدث هذا الانقسام الأول قبل أن يكون المبيض كامل التكوين (1943 ,Bacchi) وبحلول ال Metaphase للانقسام الأول فإن جدار النواة يختفي وينفصل كل زوج من الكروموزومات في ال Telophase , Anaphase ليكونا مجموعتين من الكروموزومات، وبعد فترة قصيرة في ال Interphase فإن الانقسام الثاني يفصل Split كل كروموزوم إلى اثنان ليكون هناك أربعة مجاميع متساوية من الكروموزومات وبعد الانقسام الثاني تتكون جدران الخلايا مكونة من أربعة خلايا في صف يمتد طوليا في النيوسيلة وهذه الأربعة خلايا هي الجراثيم المؤنثة.
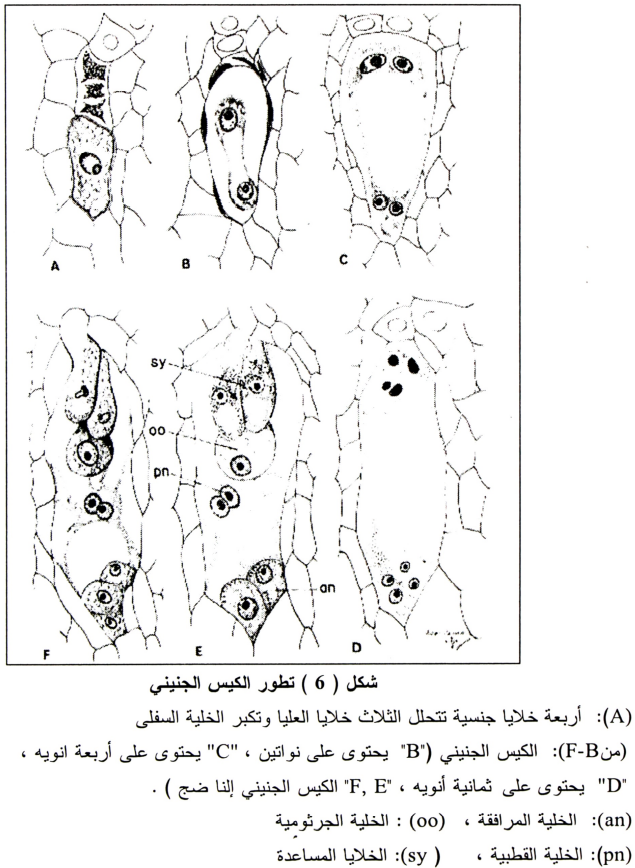
هـ- الكيس الجنيني والبويضة:
تستمر الخلية السفلى فقط في التكوين والنمو بينما تتحلل الثلاث خلايا العليا (شكل 6) وتنمو الخلية السفلى طوليا وتحتل المساحة التي خلفتها تحلل الثلاث خلايا أعلاها، وهذه الخلية الجرثومية المؤنثة الفعالة تنمو وتستمر في النمو لتكوين الكيس الجنيني وتكبر كثيرا ولكن السيتوبلازم الموجود بها لا يزيد بما يساير هذه الزيادة في الحجم، وعلى هذا تظهر فجوات كثيرة وعندما يصبح الكيس الجنيني ناضجا فإنها تحتل أكبر جزء من الحجم داخل جدران الخلايا. ومع نمو الجرثومة المؤنثة فإن النواة تنقسم وتتجه النواتين الناتجتين إلى اتجاهين مختلفين (شكل 6-B) وكل من هذه الأنوية تنقسم ثانية حتى يصبح هناك أربعة أنوية في الكيس الجنيني (شكل 6-C) وكل من هذه الأربعة أنوية تنقسم مره ثانية وبذلك تنتج ثمانية أنوية (يتواجد أربعة أنوية في كل طرف من أطراف الكيس الجنيني) (شكل 5) وتبقى ثلاثة أنوية قرب الطرف السفلى (قرب الكلازا) ومنها يتكون ثلاث خلايا قطبية Antipodal cells (شكل 6-D) وفي جهة الميكروبيل تنتظم أيضا ثلاثة أنوية أحدها البويضة والأخيرتان هما الخلايا المساعدة Synergids والتي قد تسهل عملية إخصاب البويضة (شكل 6-E)، وفي هذه المرحلة تكون البويضة ناضجة وصالحة للإخصاب وتكون النواتين الباقيتين (واحدة من كل طرف من الكيس الجنيني النواة القطبية Polar nuclei ثم تتحرك إلى وسط الكيس الجنيني (شكل 6-F) ومن المفروض إنها تلتحم لتكون نواه الأندوسبرم.
وتتكون مراحل التطور المتناظرة متأخرة في المبيض عنها في المتوك لنفس الزهرة، وعلى ذلك تكون الجراثيم المذكرة قد بدأت في تكوين حبوب اللقاح قبل أن تكون الخلية الجرثومية المؤنثة Megasporocyte قد أنهت طور ال Prophase للانقسام الأول، وعند تفتح الأزهار يكون الكيس الجنيني عادة في مرحلة الثمانية أنوية أو قد يكون في مرحلة 1، 2 أو 4 أنوية (1943 ,Bacchi)
2- تكون حبوب اللقاح والجاميطات المذكرة
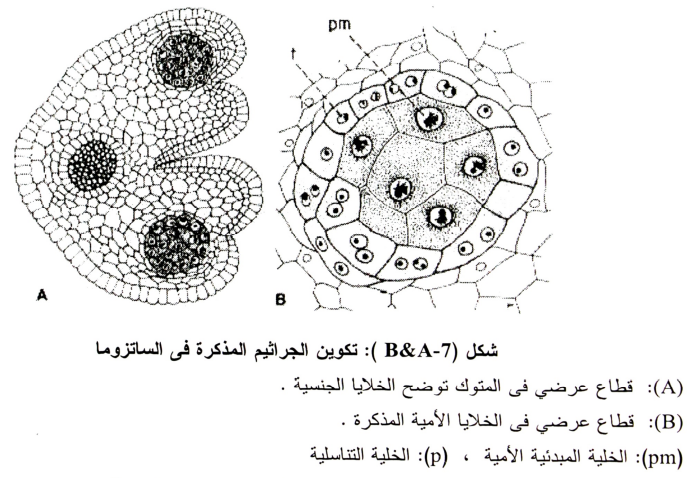
أ- تكوين الخلية الامية لحبوب اللقاح:
في المراحل المبكرة من تكوين المتوك تكون الخلية المكونة له مميزة بنواة، ومنشأ هذه الخلايا من طبقة تحت البشرة كما هو الحال في الأعضاء المؤنثة وتنقسم هذه الخلايا عرضيا (جدار عرضي) لتكون طبقة خلايا خارجية من الخلايا الأمية المغذية Parietal Cells وطبقة داخلية عبارة عن الخلايا الجرثومية، وتحدث انقسامات للخلايا الجدارية Parietal Cells وتنتج عنها أربعة طبقات من الخلايا تكون الطبقة الداخلية منهم عبارة عن النسيج المغذي Tapetum وتكون الأخرى مع البشرة جدار المتوك، والنسيج المغذي Tapetum يكون عبارة عن طبقة واحدة عادة ولكن قد يكون 2 أو 3 طبقات (1954 ,Banerji). ويحيط هذا النسيج بأسطوانة الخلايا الجرثومية (شكل7-A) والتي تتكون من الانقسامات المتتالية للخلايا الجرثومية، وقبل أن تصل الخلايا الأمية إلى منتصف ال Prophase فإن نواة كل خلية من النسيج المغذي Tapetum تنقسم مرة، كما تحدث انقسامات أخرى بعد ذلك بحيث يصبح هذا النسيج ثنائي النواة أو عديد الأنوية بدون انتظام، ومع تقدم تكوين حبوب اللقاح تتحلل خلايا هذا النسيج المغذي (قد يكون لإمداد حبوب اللقاح بالغذاء اللازم لتكوينها).
ب- الانقسام الاختزالي والجراثيم في النباتات الثنائية:
كل الطراز المنزرعة من الموالح والتي تشمل أجناس ال Poncirus, Citrus, Fortunella ثنائية (9 = X)، كما تتواجد نباتات ثنائية أيضا في Severinia، Atlantia ، Afraegle ، Murraya ، Feronia ، Aglopsis ، Citropsis ، Triphasia ,Micromlium ، Erimocitrus ، Microcitrus ، Clausena.
والخلية الأمية لحبوب اللقاح قبل الانقسام الاختزالي الأول تكون متميزة عن خلايا النسيج المغذي Tapetum والمتواجدة حولها حيث تكون ذات حجم كبير وتحتوي على نواة واحدة وذات صبغة مختلفة (شكل 7-B) وفي المرحلة المتأخرة من ال Prophase من الانقسام الأول فإن الخلية الأمية تستدير وتبقى جدران الخلية الأساسية الجرثومية الأمية في صورتها الأساسية إلى أن تنكسر في نهاية مرحلة ال Sporad والتي يكون فيها السيتوبلازم جدار جديد مستدير داخل الجدار الرئيسي بمسافة (شكل 8-A).
وفي الخلايا الأمية لحبوب اللقاح في الموالح Citrus (كما هو الحال في الخلية الأمية المؤنثة) ففي ال Prophase للانقسام الأول يتقدم التطور بصورة طبيعية حيث أن في المراحل المتقدمة تحتوي الأصناف الثنائية على تسعة کروموزومات مزدوجة Bivalent (شكل 8-B).
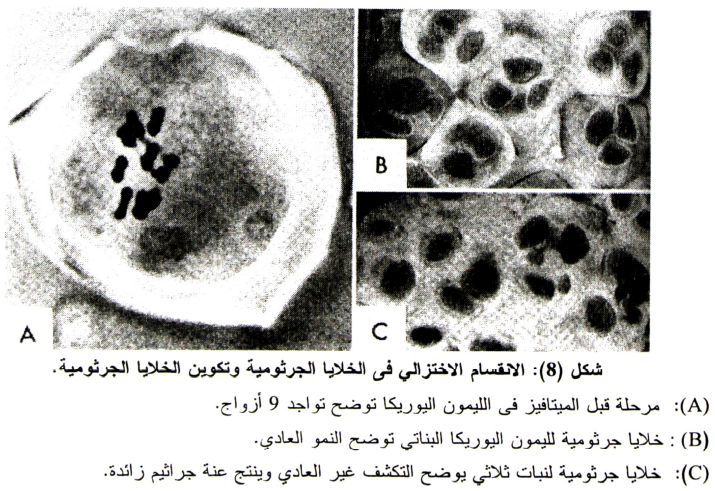
ونجد عند مرحلة ال Diakenesis أن الكروموزمات تكون متناثرة ومتباعدة في النواة (شكل 8-C) ويختفي الجدار النووي قبل ال Metaphase وعند هذه المرحلة تصبح التسع كروموزمات المتزاوجة منتظمة على الصفيحة الوسطى بين المغزل (والذي يكون قد تكون عند هذا الوقت) ويكون متوازيا مع محوره، وقد وجد بعض حالات الارتباطات الثانوية للكروموزومات أثناء بداية أول Metaphase، فقد ذكر (1966 ,Iwanasa) أنه قد وجد أذرع متعددة Multivalents، وانقلابات Inversions، وأذرع وحيدة Univalent، كما وجدت حالة انتقال (إحلال) Translocation في البرتقال الفالنشيا، وانقلاب في الليمون المكسيكي وفشل في الانفصال Asynapsis.
وفي طور (1) Anaphase تنفصل التسعة ثنائي الكروموزومات بطريقة عادية مما ينتج عنه نواتان جديدتان، وبعد مرحلة قصيرة في Interkinesis تختفي جدر النواتان المتكونتان ويبدأ الانقسام الثاني بطريقة عادية. وعادة يوجد مجموعتان من 9 کروموزومات في كل من ال Metaphase للانقسام الثاني، لكنه لوحظ عدم تساوي في التوزيع في بعض الحالات مما يدل على عدم انتظام انفصال الكروموزومات في الانقسام الأول، وقد يرجع ذلك إلى تواجد ارتباط ثانوي للكروموزومات (1954 , Banerji)، ويكون هناك في نهاية الانقسام الثاني أربعة مجاميع من 9 کروموزومات مرتبة في أربعة أنوية في داخل الجدار المستدير للخلية الأم وبذلك يكون طور الأربع أنوية قد تكون.
وتتكون الجدران بين الأربعة أنوية وتنفصل الأربعة خلايا الجديدة (الجراثيم أو حبوب اللقاح) وتستدير في داخل الجدار الأساسي للخلية الأم وهذا هو الطور الرباعي لحبوب اللقاح Pollen tetrad وحيث أنه يمكن أن تتكون أكثر من أربعة جراثيم من الخلية الأمية الواحدة (أحيانا في الثنائي "2ن"، وغالبا في متعدد المجاميع الكروموزومية) نجد أن الاصطلاح السابق قد يكون غير مناسب ويفضل استخدام لفظ Sporad أو Webber, 1933) Microsporad).
ج- الانقسام الاختزالي والجراثيم في عديدة المجاميع الكروموزومية:
يؤدي تواجد مجموعة كروموزومية إضافية (3ن) أو مجموعتين (4ن) إلى عدة خواص مختلفة عن الأشجار الثنائية سواء كان ذلك بالنسبة للأشجار أو الثمار في حالة الأشجار الثلاثية (3ن) نجد أن اختزال الكروموزومات لا يمكن أن يكون منتظما حيث أن هناك ثلاث مجاميع من الكروموزومات من كل نوع, ولذا يصعب فصل الكروموزومات في الانقسام الأول لكل نوع إلى مجموعتين، كما وجد أنه في الأشجار الرباعية (ذات التضاعف الذاتي) يكثر تواجد تعدد الأفرع Multivalents بين الكروموزومات في الانقسام الاختزالي مما ينتج عنه انقسام غير منتظم (.Russo and Torrisi, 1951 b).
د- تطور حبه اللقاح والخلية الجرثومية:
يتبع تطور وتكوين حبه اللقاح الطريقة الشائعة لمغطاة البذور حيث تكبر الجرثومة ويتكون فيها جدارين سميكين خارجي (Exine) وداخلي (Intine)، وقبل انتثار المتوك تنقسم النواة مكونة النوية الخضرية والنواة الجرثومية. ويكون لون المتوك عادة أصفر ناصع عند النضج، بينما يكون لون المتوك أخف من ذلك في الأصناف التي تكون حبوب لقاحها بها عيوب تكوينية شديدة، بينما لون المتوك في الحالات التي لا تنتج حبوب لقاح بالمرة يكون باهتا كريمي أو أبيض. وقد استخدمت المظاهر المورفولوجية لحبة اللقاح للتمييز بين الأنواع المختلفة سواء عن طريق متوسط القطر (من 34-36 U) أو في النظام الشبكي الموجود على سطح حبه اللقاح.
 الاكثر قراءة في مقالات منوعة عن الحمضيات
الاكثر قراءة في مقالات منوعة عن الحمضيات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام) قسم الشؤون الفكرية يصدر مجموعة قصصية بعنوان (قلوب بلا مأوى)
قسم الشؤون الفكرية يصدر مجموعة قصصية بعنوان (قلوب بلا مأوى) قسم الشؤون الفكرية يصدر مجموعة قصصية بعنوان (قلوب بلا مأوى)
قسم الشؤون الفكرية يصدر مجموعة قصصية بعنوان (قلوب بلا مأوى)



















