























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Elongation Factors Bind Alternately to the Ribosome
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 25-5-2021
25-5-2021
 2656
2656




+
-
20
Elongation Factors Bind Alternately to the Ribosome
KEY CONCEPTS
- Translocation requires EF-G, whose structure resembles the aminoacyl-tRNA–EF-Tu–GTP complex.
- Binding of EF-Tu and EF-G to the ribosome is mutually exclusive.
- Translocation requires GTP hydrolysis, which triggers a change in EF-G, which, in turn, triggers a change in ribosome structure.
Translocation requires GTP and another elongation factor, EF-G. (The eukaryotic homolog of EF-G is eEF2.) This factor is a major constituent of the cell; it is present at a level of about 1 copy per ribosome (20,000 molecules per cell).
Ribosomes cannot bind EF-Tu and EF-G simultaneously, so translation follows the cycle illustrated in Figure 1, in which the factors are alternately bound to and released from the ribosome. Thus, EF-Tu–GDP must be released before EF-G can bind, and then EF-G must be released before aminoacyl-tRNA–EF-Tu–GTP can bind.
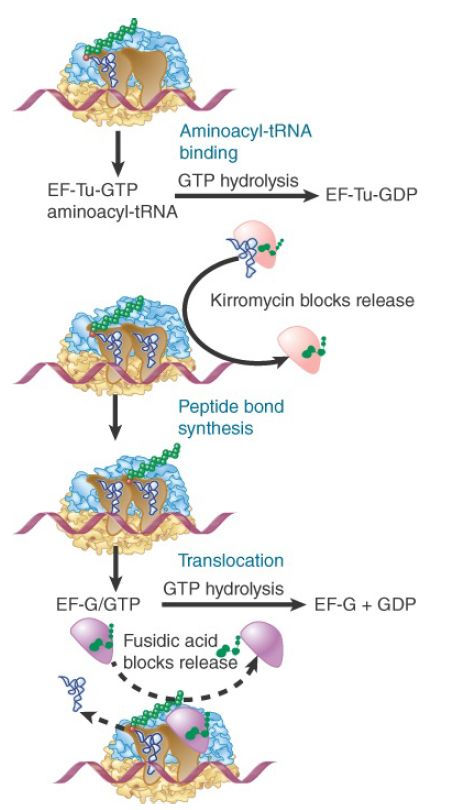
FIGURE 1. Binding of factors EF-Tu and EF-G alternates as ribosomes accept new aminoacyl-tRNAs, form peptide bonds, and translocate.
Does the ability of each elongation factor to exclude the other rely on an allosteric effect on the overall conformation of the ribosome or on direct competition for overlapping binding sites? Figure 2 shows an extraordinary similarity between the structures of the ternary complex of aminoacyl-tRNA–EF-Tu–GDP and EF-G. The structure of EF-G mimics the overall structure of EF-Tu bound to the amino acceptor stem of aminoacyl-tRNA. This suggests that they compete for the same binding site (presumably in the vicinity of the A site). The need for each factor to be released before the other can bind ensures that the events of translation proceed in an orderly manner.
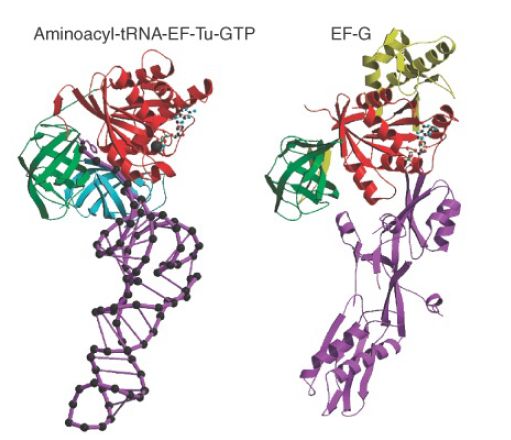
FIGURE 2. The structure of the ternary complex of aminoacyltRNA–EF-Tu–GTP (left) resembles the structure of EF-G (right). Structurally conserved domains of EF-Tu and EF-G are in red and green; the tRNA and the domain resembling it in EF-G are in purple.
Photo courtesy of Poul Nissen, University of Aarhus, Denmark.
Both elongation factors are monomeric GTP-binding proteins that are active when bound to GTP but inactive when bound to GDP. The triphosphate form is required for binding to the ribosome, which ensures that each factor obtains access to the ribosome only in the company of the GTP that it needs to fulfill its function. EF-G binds to the ribosome to facilitate translocation and then is released following ribosome movement. EF-G can still bind to the ribosome when GMP-PCP is substituted for GTP, so the presence of a guanine nucleotide is needed for binding, but its hydrolysis is not absolutely essential for translocation (though translocation is much slower in the absence of GTP hydrolysis). The hydrolysis of GTP is needed to release EF-G.
The need for EF-G release was discovered by the effects of the steroid antibiotic fusidic acid, which “jams” the ribosome in its posttranslocation state. In the presence of fusidic acid, one round of translocation occurs; EF-G binds to the ribosome, GTP is hydrolyzed, and the ribosome moves over by three nucleotides. However, fusidic acid stabilizes the ribosome–EF-G–GDP complex so that EF-G and GDP remain on the ribosome instead of being released. As a result, the ribosome cannot bind aminoacyl-tRNA, and no further amino acids can be added to the chain.
Translocation is an intrinsic property of the ribosome that requires a major change in structure (see the section later in this chapter titled Ribosomes Have Several Active Centers). This intrinsic translocation is activated by EF-G in conjunction with GTP hydrolysis, which occurs before translocation and accelerates the ribosomal movement. The most likely mechanism is that GTP hydrolysis causes a change in the structure of EF-G, which, in turn, forces a change in the ribosome structure. An extensive reorientation of EF-G occurs at translocation. Before translocation, it is bound across the two ribosomal subunits. Most of its contacts with the 30S subunit are made by a region called domain 4, which is inserted into the A site. This domain could be responsible for displacing the tRNA. After translocation, domain 4 is instead oriented toward the 50S subunit.
The eukaryotic counterpart to EF-G is the protein eEF2, which functions in a similar manner to a translocase dependent on GTP hydrolysis. Its action also is inhibited by fusidic acid. A stable
complex of eEF2 with GTP can be isolated and the complex can bind to ribosomes with consequent hydrolysis of its GTP. A unique property of eEF2 is its susceptibility to diphtheria toxin.
The toxin uses nicotinamide adenine dinucleotide (NAD) as a cofactor to transfer an adenosine diphosphate ribosyl (ADPR) moiety onto the eEF2. The ADPR–eEF2 conjugate is inactive in translation. The substrate for the attachment is an unusual amino acid that is produced by modifying a histidine; it is common to the eEF2 of many species.
The ADP-ribosylation is responsible for the lethal effects of diphtheria toxin. The reaction is extremely effective: A single molecule of toxin can modify enough eEF2 molecules to kill a cell.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















