























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Specific Sequences Attach DNA to an Interphase Matrix
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 22-3-2021
22-3-2021
 2246
2246




+
-
20
Specific Sequences Attach DNA to an Interphase Matrix
KEY CONCEPTS
-DNA is attached to the nuclear matrix at sequences called matrix attachment regions.
-The matrix attachment regions on average are A-T rich but do not have any specific consensus sequence.
Is DNA attached to a matrix via specific sequences? Researchers can empirically define DNA sites attached to proteinaceous structures in interphase nuclei. They are called matrix attachment regions (MARs) or scaffold attachment regions (SARs). The precise functionality of the nuclear matrix and MARs has been a topic of considerable debate. Some observations are clear: The same sequences appear to attach to the protein substructure in both metaphase and interphase cells. Chromatin appears to be attached to an underlying structure in vivo, and there have been many suggestions that this attachment affects aspects of transcription, repair, or replication.
Are particular DNA regions associated with this matrix? FIGURE 1 summarizes two approaches to detect specific MARs. Both begin by isolating the matrix as a crude nuclear preparation containing chromatin and nuclear proteins. Researchers can then use different treatments to characterize DNA in the matrix or to identify DNA able to attach to it. The same general approaches can be applied to metaphase scaffold preparations.
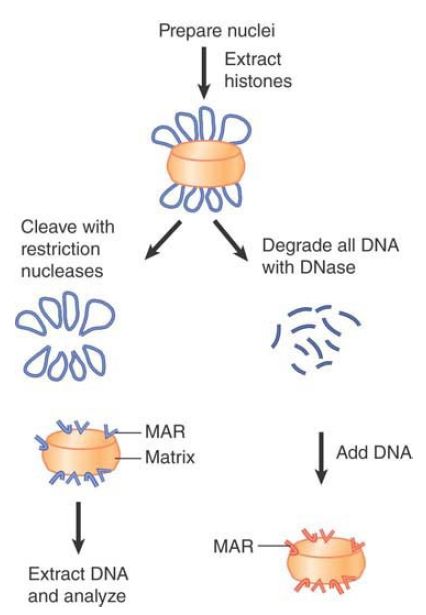
FIGURE 1 MARs can be identified by characterizing the DNA retained by the matrix isolated in vivo (left) or by identifying the fragments that can bind to the matrix from which all DNA has been removed (right).
To analyze existing MARs that are bound to the matrix in vivo, chromosomal loops can be decondensed by extracting the chromatin proteins. Removal of the DNA loops by treatment with restriction nucleases leaves only the (presumptive) in vivo MAR sequences attached to the matrix.
The complementary approach is to remove all of the DNA from the matrix by treatment with DNase, at which point isolated fragments of DNA can be tested for their ability to bind to the matrix in vitro.
The same sequences should be associated with the matrix in vivo or in vitro. After researchers identify a potential MAR, they can determine the size of the minimal region needed for association in vitro by deletions, aiding in the identification of MAR-sequencebinding proteins.
A surprising feature is the lack of conservation of sequence in MAR fragments. Other than A-T richness, they lack any other obvious consensus sequences. Other interesting sequences, however, often are in the DNA stretch containing the MAR. cis-acting sites that regulate transcription are common, as are 5′ introns and recognition sites for topoisomerase II. It is therefore possible that a MAR serves more than one function by providing a site for attachment to the matrix and containing other sites at which topological changes in DNA are effected.
What is the relationship between the chromosome scaffold of dividing cells and the matrix of interphase cells? Are the same DNA sequences attached to both structures? In several cases, the same DNA fragments that are found within the nuclear matrix in vivo can be retrieved from the metaphase scaffold. Fragments that contain MAR sequences can bind to a metaphase scaffold, so it therefore seems likely that DNA contains a single type of attachment site. In interphase cells the attachment site is connected to the nuclear matrix, whereas in mitotic cells it is connected to the chromosome scaffold. Interestingly, it is also clear that although some MARs are constitutive (continuously bound to the matrix or scaffold), others appear to be facultative and change their interactions with the matrix depending on cell type or other conditions.
The nuclear matrix and chromosome scaffold consist of different proteins, although there are some common components. Topoisomerase II is a prominent component of the chromosome
scaffold, and is a constituent of the nuclear matrix, reflecting the importance of topology in both cases.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















